|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Функциональная организация базальных ганглиев
Как и мозжечок, они не имеют непосредственного эфферентного выхода к спинному мозгу и, так же как он, не могут напрямую контролировать движения. В отличие от мозжечка, они не получают непосредственную информацию от различных рецепторов, но используют сигналы, поступающие от многочисленных областей коры. Их эфферентная связь направлена через таламус к префронтальной, вторичной и первичной моторной коре: базальные ганглии модулируют активность этих двигательных центров.
Все афферентные связи базальных ганглиев приходятся на полосатое тело. В нём оканчиваются многочисленные глутаматэргические волокна, берущие своё начало от нейронов сенсомоторных и ассоциативных областей коры, планирующих движение и создающих двигательные программы. Источником для другой группы сигналов, поступающих к полосатому телу, служит таламус. Ещё один важный афферентный путь начинается в дорсальной части чёрной субстанции: её дофаминэргические нейроны оказывают тормозное действие на полосатое тело, в котором количество дофаминэргических синапсов на единицу объёма больше, чем в любой другой области мозга. В полосатом теле есть нейроны локальных сетей, перерабатывающие полученную информацию, и выходные нейроны, передающие сигналы двум получателям: бледному шару и ретикулярной части чёрной субстанции. Свыше 90% выходящих из полосатого тела нейронов используют тормозной медиатор – ГАМК, остальные – ацетилхолин, субстанцию Р, соматостатин и энкефалин, которые оказывают возбуждающее или модулирующее действие. Выходная активность ретикулярной части чёрной субстанции и бледного шара адресована таламусу. Получившие возбуждение нейроны таламуса распределяют его между двумя получателями: во-первых, они передают информацию префронтальной ассоциативной области, вторичной и первичной моторной коре, а во-вторых, с помощью холинэргических нейронов отправляют её полосатому телу. В результате формируются два круга, по которым может циркулировать возбуждение: большой и малый (рис. 10.12). По большому кругу возбуждение от различных областей коры, в том числе от моторных, поступает в полосатое тело, оттуда в бледный шар или ретикулярную часть чёрной субстанции, затем в таламус, а из него возвращается в моторную кору. В малом круге оно циркулирует по маршруту: полосатое тело – бледный шар или чёрная субстанция – таламус – полосатое тело.
Связи базальных ганглиев с корой организованы соматотопически, разные круги переключения предусматривают отдельную обработку информации для движений ног, рук, глаз, а также комплексов действий, необходимых для когнитивных форм поведения. Так, например, кортикоспинальный путь, начинающийся от первичной и вторичной моторной, соматосенсорной коры и префронтальной области снабжает моторную часть скорлупы многочисленными и хорошо топографически организованными входами, при этом большая часть волокон поступает к базальным ганглиям от добавочного моторного ареала и премоторной области. К моторным областям проецируется через таламус и выходная активность базальных ганглиев, что позволяет им участвовать в управлении важнейшими нисходящими моторными путями. Активность некоторых нейронов базальных ганглиев похожа на деятельность клеток моторной коры: отдельные нейроны активируются только при изменении положения конкретного сустава в определённом направлении. В то же время не обнаруживается связи между их активностью и силой сокращения отдельных мышц. Электрическая активность базальных ганглиев регистрируется непосредственно перед началом произвольных движений контрлатеральных конечностей. По-видимому, базальные ганглии участвуют в переработке информации, необходимой для планирования и начальной стадии произвольных движений, а также в формировании необходимой для выполнения движения позы. Наличие в составе базальных ганглиев возбуждающих и тормозных нейронов позволяет сбалансировать их выходную активность таким образом, чтобы обеспечить нужную плавность движений. Считают, что базальные ганглии облегчают одни и угнетают другие движения подобно тому, как в сенсорных системах к разным последствиям приводит стимуляция центральной части рецептивного поля и его периферии. Возможно, что такая деятельность базальных ганглиев относится к чередованию тонических и фазных компонентов движения, т.е. формированию позы, требующейся для совершения конкретного движения, а затем и самого движения. Сопоставление функций мозжечка и базальных ганглиев в управлении движением позволяет найти несколько различий между ними. Во-первых, мозжечок получает входы и от сенсомоторной коры, и от периферии, а базальные ганглии только от коры, но не от одних лишь моторных областей, а и от ассоциативных. Во-вторых, мозжечок посылает эфферентные сигналы только к первичной и вторичной моторной коре, а базальные ганглии, наряду с этими областями, имеют выход к префронтальной ассоциативной коре. В третьих, мозжечок получает информацию непосредственно от спинного мозга, а его обширные связи с двигательными ядрами ствола позволяют ему оперативно вмешиваться в управление уже совершающихся движений. У базальных ганглиев мало контактов со стволом и есть лишь непрямые связи со спинным мозгом. Эти параллели позволяют констатировать возможность управления ходом движений у мозжечка, а у базальных ганглиев – участие в стратегических аспектах движения, связанных с их планированием и инициацией. Дата добавления: 2015-12-15 | Просмотры: 913 | Нарушение авторских прав |
 Базальные ганглии представляют собой сложно организованный комплекс разнородных по происхождению ядер, которые образуют цепи модулирующих переключений (рис. 10.10).
Базальные ганглии представляют собой сложно организованный комплекс разнородных по происхождению ядер, которые образуют цепи модулирующих переключений (рис. 10.10).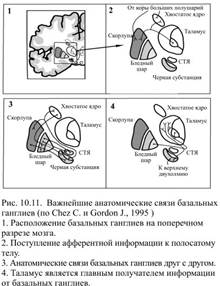 К системе базальных ганглиев относят пять пар функционально объединённых ядер: 1) хвостатое ядро, напоминающее вытянутую грушу и расположенное кпереди и кнаружи от зрительного бугра; 2) скорлупу, происходящую, как и хвостатое ядро из переднего мозга и образованную такими же, как и оно, клетками (на основании сходства обоих ядер их часто объединяют под названием полосатое тело (лат. corpus striatum) или новое полосатое тело (лат. neostriatum); 3) бледный шар (лат. globus pallidus, pallidum), происходящий из диэнцефальной (промежуточной) части мозга и разделяющийся на внутренний и внешний сегменты; 4) субталамическое ядро, находящееся книзу от таламуса на границе со средним мозгом; 5) чёрная субстанция, расположенная в среднем мозгу и состоящая из двух частей: вентральной, бледной на вид и по составу клеток сходной с бледным шаром, и дорсальной – сильно пигментированной, что и послужило поводом для названия этой структуры чёрной – этот цвет обусловлен присутствием нейромеланина, являющегося синтетическим предшественником дофамина. Две части чёрной субстанции находятся в реципрокных отношениях друг с другом. Бледный шар и отделённая от него внутренней капсулой вентральная часть чёрной субстанции, которая называется ещё ретикулярной, похожи не только внешне, но и функционально: это важнейшие выходные области базальных ганглиев. Так же близки структурно и функционально хвостатое ядро и скорлупа, их обычно объединяют, как полосатое тело – к нему поступают входные сигналы от обширных областей коры (рис. 10.11).
К системе базальных ганглиев относят пять пар функционально объединённых ядер: 1) хвостатое ядро, напоминающее вытянутую грушу и расположенное кпереди и кнаружи от зрительного бугра; 2) скорлупу, происходящую, как и хвостатое ядро из переднего мозга и образованную такими же, как и оно, клетками (на основании сходства обоих ядер их часто объединяют под названием полосатое тело (лат. corpus striatum) или новое полосатое тело (лат. neostriatum); 3) бледный шар (лат. globus pallidus, pallidum), происходящий из диэнцефальной (промежуточной) части мозга и разделяющийся на внутренний и внешний сегменты; 4) субталамическое ядро, находящееся книзу от таламуса на границе со средним мозгом; 5) чёрная субстанция, расположенная в среднем мозгу и состоящая из двух частей: вентральной, бледной на вид и по составу клеток сходной с бледным шаром, и дорсальной – сильно пигментированной, что и послужило поводом для названия этой структуры чёрной – этот цвет обусловлен присутствием нейромеланина, являющегося синтетическим предшественником дофамина. Две части чёрной субстанции находятся в реципрокных отношениях друг с другом. Бледный шар и отделённая от него внутренней капсулой вентральная часть чёрной субстанции, которая называется ещё ретикулярной, похожи не только внешне, но и функционально: это важнейшие выходные области базальных ганглиев. Так же близки структурно и функционально хвостатое ядро и скорлупа, их обычно объединяют, как полосатое тело – к нему поступают входные сигналы от обширных областей коры (рис. 10.11).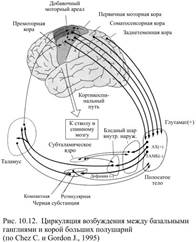 Субталамическое ядро получает сигналы от бледного шара и от моторной коры, а выходную активность адресует двум отделам бледного шара: латеральному и медиальному, действуя на них противоположно, т.е., возбуждая один и тормозя другой. Субталамическое ядро состоит с бледным шаром в реципрокных отношениях и его роль состоит в том, чтобы контролировать выходную активность бледного шара.
Субталамическое ядро получает сигналы от бледного шара и от моторной коры, а выходную активность адресует двум отделам бледного шара: латеральному и медиальному, действуя на них противоположно, т.е., возбуждая один и тормозя другой. Субталамическое ядро состоит с бледным шаром в реципрокных отношениях и его роль состоит в том, чтобы контролировать выходную активность бледного шара.