|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
МЕХАНИЗМ ДЕЙСТВИЯ ГОРМОНОВ
Ключевым этапом физиологического действия гормона на клетку является его спонтанное и обратимое комплексообразование со специфическим белком-рецептором. Рецептор является необходимым периферическим звеном эндокринной системы. Рецепторная молекула состоит из трех главных пространственно разобщенных локусов, осуществляющих основные динамически сопряженные функции:
Основные свойства рецепторов:
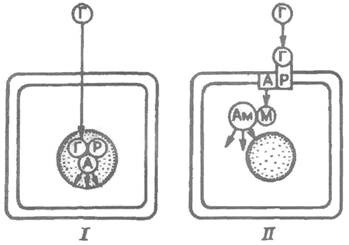
К настоящему времени идентифицировано около 60 гормональных рецепторов, причем более половины из них локализуется на мембранах клеток. Другие гормоны взаимодействуют с рецепторами, расположенными внутриклеточно (цитоплазматические, ядерные и митохондриальные). Мембранный рецептор взаимодействует с гормоном, который плохо проникает в клетку, в результате сразу же образуются внутриклеточные медиаторы. При этом механизме действия гормонов эффекты развиваются быстро. Во втором случае гормон свободно проникает в клетку через мембрану, взаимодействует с ядерными, цитозольными и т.д. рецепторами и на первых этапах своего действия не требует образования внутриклеточных медиаторов. Эти эффекты медленно развиваются и связаны с биосинтезом белка. Через поверхностные (мембранные) рецепторы действуют гормоны белковой, полипептидной природы, а также и производные аминокислот. Через внутриклеточные – стероиды и тиреоидные гормоны (рис. 3.1).
Рис. 3.1. Типы рецепции гормонов клетками. I – внутриклеточный; II – мембранный: Г – гормон; А – акцептор; М – внутриклеточный медиатор; Ам – «акцептор» медиатора; Р – рецептор.
I. Гормоны, взаимодействующие с мембранными рецепторами. А. Белковые гормоны. Гликопротеидные гормоны: ТТГ, ФСГ, ЛГ, хорионический гонадотропин, СТГ, пролактин, хорионический соматомаммотропин (плацентарный лактоген), инсулин, инсулиноподобные факторы роста I и II, соматомедины, релаксин, фактор роста нервов, гастрин, холецистокинин. Глюкагон (панкреатический и кишечный), секретин, вазоактивный кишечный пептид, желудочный ингибиторный полипептид, АКТГ и α-МСГ. Энкефалины, эндорфины, β-липотропин, окситоцин, вазопрессин. Эпидермальный фактор роста, урогастрон, паратгормон, кальцитонин. Тиреолиберин, гонадолиберин, соматостатин, соматолиберин. Б. Катехоламины. В. Простагландины. Г. Серотонин, нейротензин, вещество Р.
II. Гормоны, взаимодействующие с внутриклеточными рецепторами. А. Стероидные гормоны. Глюкокортикоиды и минералокортикоиды, эстрогены, андрогены, прогестины. Б. Тиреоидные гормоны. Ко многим гормонам рецепторы еще не идентифицированы. Так, из 30 известных простагландинов, простациклинов, тромбоксанов рецепторы определены лишь к шести. Биологический эффект гормонов, взаимодействующих с мембранными рецепторами, осуществляется с участием вторичного мессенджера (передатчика, или посредника). Различают 5 типов вторичных мессенджеров: 1. циклический аденилатмонофосфат (цАМФ) – активирует цАМФ-зависимую протеинкиназу; 2. циклический гуанозинмонофосфат (цГМФ) – активирует цГМФ-зависимую протеинкиназу; 3. ионы кальция – активируют кальмодулин и другие кальций-связывающие белки; 4. инозитол-3-фосфат – стимулирует выход Са++ из ЭПС; 5. диацилглицерол – стимулирует протеинкиназу С.
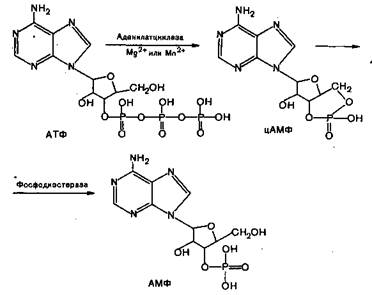
1. При изучении механизма действия глюкагона и адреналина на процессы распада гликогена в печени Сатерлендом и др. (1959) установлено, что влияние этих гормонов осуществляется посредством стимуляции активности фермента клеточной мембраны аденилатциклазы, который катализирует превращение внутриклеточного аденозинтрифосфата (АТФ) в циклический аденилатмонофосфат (цАМФ). За это открытие Сатерленд удостоен Нобелевской премии 1971 года. Аденилатциклаза – мембранный белок, связанный с рецептором, регуляторным белком (Gs), которые в обычном (т.е. нестимулированном) состоянии разобщены между собой (рис. 3.2). Рецептор располагается на внешней, а аденилатциклаза - на внутренней поверхности плазматической мембраны.
Связывание первичного мессенджера (гормона) с рецептором ведет к его конформационным изменениям, следствием чего является стимуляция Gs регуляторного белка, который активирует аденилатциклазу. Активированная аденилатциклаза катализирует превращение цитоплазматического АТФ в циклический аденозинмонофосфат (цАМФ). Концентрация цАМФ в клетке увеличивается через 1-5 минут после действия гормона с 10-8 до 10-6 М. Вторичный мессенджер – цАМФ – в свою очередь стимулирует протеинкиназы, представляющие собой ферменты, которые фосфорилируют соответствующие белки (часто энзимы), которые осуществляют физиологический ответ (секреция, сокращение и т.д.). После этого цАМФ под влиянием фосфодиэстеразы превращается в неактивную форму. Таким образом, активация аденилатциклазы регуляторным белком Gs запускает каскад реакций, приводящих к физиологическому ответу.
Рис. 3.2. Роль вторичного мессенджера цАМФ в механизмах действия гормонов.
Такой каскад работает как биохимический усилитель. Действительно, одна активная молекула активной аденилатциклазы катализирует превращение100 молекул цАМФ, которые в свою очередь катализируют превращение 10 000 молекул фосфорилированных белков и 1 000 000 молекул конечных продуктов реакции, приводящих к физиологическому ответу.
Таблица 3.1. Дата добавления: 2015-02-06 | Просмотры: 1316 | Нарушение авторских прав |