|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
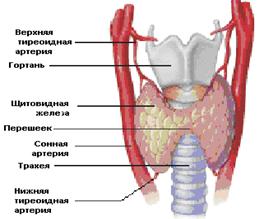
ЩИТОВИДНАЯ ЖЕЛЕЗАЩитовидная железа (ЩЖ) - небольшой орган, имеет форму бабочки и располагается на шее спереди от трахеи и снизу от гортани (рис. 7.1). В норме масса щитовидной железы составляет от 20 до 60 г, размеры долей варьируют в пределах 5-8х2-4х1-3см. В период полового созревания масса щитовидная железа увеличивается, а в старческом возрасте уменьшается. У женщин щитовидная железа больше, чем у мужчин; во время беременности происходит ее физиологическое увеличение, которое исчезает самостоятельно в течение 6—12 мес. после родов. Она состоит из двух долей, соединенных перешейком.
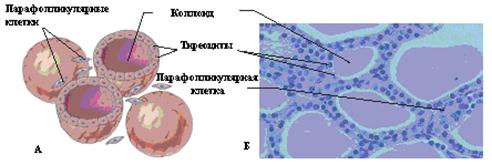
Рис. 7.1. Расположение щитовидной железы. Щитовидная железа является одним из наиболее кровоснабжаемых органов с развитой артериальной и более мощной венозной системами (величина кровотока в ЩЖ на 0,1кг ее массы составляет 560 мл/мин, в почке – 420 мл/мин, в печени – 150 мл/мин, в мозге – 65 мл/мин). Кровь в железу поступает по двум верхним тиреоидным артериям (ветви наружной сонной артерии) и двум нижним тиреоидным артериям, которые образуют между собой анастомозы. Венозная и лимфатическая системы осуществляют отток от щитовидной железы крови и лимфы, содержащих тиреоидные гормоны, тиреоглобулин, а при патологических состояниях антитиреоидные антитела, тиреостимулирующие и тиреоблокирующие иммуноглобулины. Иннервация щитовидной железой осуществляется веточками как блуждающего нерва (парасимпатическая), так и ветвями шейных ганглиев (симпатическая). Структурной и функциональной единицей щитовидной железы являются фолликулы — пузырьки различной формы, чаще округлой, диаметром 25 - 500 мкм, отделенные друг от друга тонкими прослойками рыхлой соединительной ткани с большим количеством кровеносных и лимфатических капилляров (рис 7.2).
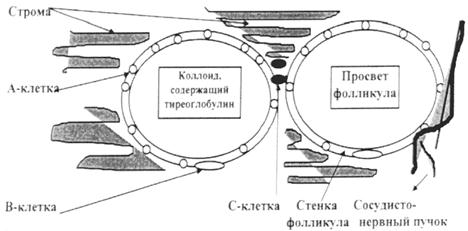
Рис. 7.2. Гистология щитовидной железы: А – схематическое строение фолликулов; Б – микроскопический препарат. Строма фолликулов заполнена коллоидом — бесструктурной массой, содержащей тиреоглобулин, который синтезируется фолликулярными, или так называемыми А-клетками, тиреоцитами, формирующими стенку фолликула. В норме они имеют кубическую форму, при повышении функциональной активности – цилиндрическую, а при снижении тиреоидной функции они уплощаются. В тиреоцитах синтезируется 2 йодсодержащих гормона – тироксин (Т4) и трийодтиронин (Т3). Последние получили свои названия по количеству атомов йода в их молекулах. Тироксин и трийодтиронин синтезируются в апикальной части тиреоидного эпителия и частично в интрафолликулярном пространстве, где накапливаются, включаясь в состав тиреоглобулина. Кроме того, в щитовидной железе имеются межфолликулярные островки эпителиальных клеток (В-клетки) (рис. 7.3), являющихся источником образования новых фолликулов. Последние, расположенные на наружной поверхности фолликулов, являются нейроэндокринными клетками, не поглощают йод и относятся к АПУД-системе; в них синтезируется гормон кальцитонин.
Рис. 7.3. Схема строения щитовидной железы.
Регуляция деятельности щитовидной железы
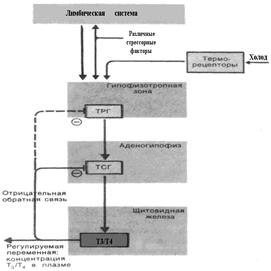
Функция ЩЖ находится под контролем гипоталамо-гипофизарной системы. Секреция регулируется тропным гормоном – ТТГ, который в свою очередь выделяется со скоростью, зависящей от активности рилизинг-гормона ТРГ. Латентный период увеличения секреции весьма мал. Согласно современным представлениям, ТТГ действует на мембраны эпителиальных клеток щитовидной железы, вызывая увеличение синтеза гормонов и их секреции путем стимуляции активности аденилатциклазы. Отрицательная обратная связь. При снижении Т3 иТ4 содержание ТТГ в плазме заметно повышается; напротив, концентрацию ТТГ можно снизить введением Т3 и Т4. Следовательно, существует механизм отрицательной обратной связи. Результаты опытов с микроинъекциями показывают, что рецепторные элементы, реагирующие на Т3 и Т4 и опосредующие отрицательную обратную связь, расположены в аденогипофизе. Остается неясным, оказывает ли цепь отрицательной обратной связи, регулирующая уровень Т3 и Т4, влияние на выделение рилизинг-гормона (ТРГ) в гипофизотропной зоне гипоталамуса (пунктирная линия на рис. 7.4).
Рис. 7.4. Схема регуляции уровня тиреоидных гормонов. «Рецепторы» Т3 и Т4 в аденогипофизе опосредуют отрицательную обратную связь.
На секрецию Т3 и Т4 оказывают влияние как внутренние, так и наружные (рис. 7.4) терморецепторы. Проведенные недавно исследования показывают, что усиление образования Т3 и Т4, вызванное холодом, значительно менее выражено у человека и других приматов, нежели у ранее изученных лабораторных животных. В любом случае при длительном воздействии холода образование гормона постепенно возвращается к начальному уровню. Таким образом, состояние акклиматизации к холоду совсем не эквивалентно гипертиреозу; чувствительность к норадреналину системы кровообращения практически одинакова у животных, адаптированных к холоду и теплу. Влияние стресс-факторов на секрецию. Помимо холода многие другие стимулы (стресс-факторы) вызывают увеличение секреции тиреоидных гормонов. Однако и в этих случаях, как при воздействии острого холодового стресса и под влиянием акклиматизации к холоду, концентрация гормонов в плазме почти не изменяется. Этот эффект отличается от ситуации с глюкокортикоидами тем, что изменяется скорость обновления, а не заданное значение и секреция. При стрессе происходит активация тиреоидного механизма (тиреоидной оси): неокортекс → септально-гиппокампово-гипоталамическое возбуждение → тиролиберин гипоталамуса → ТТГ аденогипофиза → тиреоидные гормоны щитовидной железы → трийодтиронин (Т3) и тироксин (Т4). Тиреоидные гормоны повышают чувствительность тканей к циркулирующим в крови катехоламинам, повышают уровень энергообразования, активизируют деятельность сердца (ЧСС, сократимость), вызывают повышение артериального давления. Синтез йодсодержащих гормонов Синтез йодсодержащих тиреоидных гормонов происходит в тиреоцитах из аминокислоты тирозина. Аденомеры щитовидной железы являются замкнутыми фолликулами, поэтому секрет, выделяемый тиреоцитами в просвет аденомера, может попасть в циркуляцию, только вновь пройдя через стенку фолликула в центробежном направлении. Поэтому в секреторном цикле щитовидной железы, в отличие от прочих желез, к фазам продукции (биосинтеза) секрета и его выделения в просвет аденомера добавляется еще дополнительная фаза – фаза выведения продукта, накопления в полости фолликула, в прифолликулярные сосуды. В выработке секреторного продукта щитовидной железы – тиреоглобулина – различают два относительно самостоятельных процесса: 1) образование белкового (гликопротеидного) компонента будущего тиреоглобулина; 2) включение в этот белковый субстрат йода. Синтез исходного гликопротеида происходит в эндоплазматической сети тиреоцитов и образующийся здесь преколлоид выделяется в полость фолликула. Одновременно с синтезом тиреоглобулина протекает окисление йодида (рис. 7.5). Этот процесс можно разделить на четыре стадии. 1. Включение йода в щитовидную железу. Йод в виде органических и неорганических соединений поступает в желудочно-кишечный тракт с пищей и питьевой водой. Йод всасывается в кишечнике в форме йодидов. С током крови они достигают щитовидной железы, ткань которой обладает уникальной способностью захватывать и концентрировать йодид со скоростью около 2 мкг в час. Транспорт йодида через мембрану тиреоцита является активным, требующим энергии процессом, при котором йодид поступает из среды с меньшей концентрацией (плазма крови) в среду с высокой концентрацией (ткань щитовидной железы).
Рис. 7.5. Последовательность фаз секреции йодсодержащих гормонов в тиреоцитах.
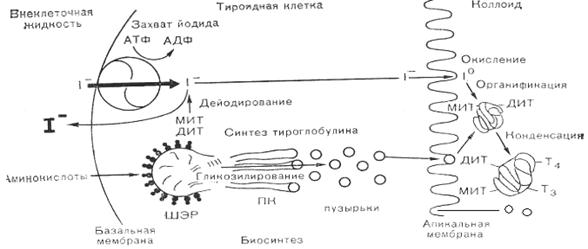
2. Органификация йода. Следующим этапом после захвата йода щитовидной железой является синтез тиреоидных гормонов. Поступивший в щитовидную железу йодид окисляется до активной формы при помощи фермента йодпероксидазы (пероксидазы) и перекиси водорода (Н2О2). Пероксидаза (мол. м. 64 000) непосредственно связана с мембраной тиреоцита. Активированный таким образом йодид (I+) способен йодировать молекулу тирозина с образованием монойодтирозина (МИТ) или дийодтирозина (ДИТ). При помощи этой пероксидазной системы щитовидная железа использует каждый поступающий в нее атом йода и препятствует возможному возвращению йодида в кровяное русло. Процесс образования МИТ и ДИТ происходит на боковых цепях молекулы тиреоглобулина, несущих тирозиновые остатки. Йодирование тиреоглобулина осуществляется на границе между апикальной частью тироцита и коллоидом, содержащимся в просвете фолликулов (рис. 7.6).
Рис. 7.6. Схема биосинтеза тиреоидных гормонов в клетке. Т3 и Т4 образуется и хранится связанными с тиреоглобулином в коллоиде. ПК – пластинчатый комплекс; МИТ – монойодтирозин; ДИТ − дийодтирозин; ШЭР – шероховатый эндоплазматический ретикулум.
3. Процесс конденсации. В заключительной стадии гормоносинтеза МИТ и ДИТ под влиянием окислительных ферментов конденсируются с образованием биологически активных тиреоидных гормонов: трийодтиронина – Т3 и тироксина – Т4. При конденсации двух субъединиц ДИТ образуется Т4; если происходит конденсирование между субъединицами ДИТ и МИТ, образуется Т3. Бóльшая часть фолликулов щитовидной железы предназначена для хранения тиреоглобулина, и количество тиреоидных гормонов таково, что если полностью блокировать биосинтез гормонов, то запасов Т3 и Т4 будет вполне достаточно для поддержания эутиреоидного состояния более месяца. 4. Высвобождение гормонов щитовидной железы. Тиреоглобулин вследствие большой величины своих молекул не может диффундировать через эпителий фолликулярной стенки. Свободное излияние коллоида из фолликула в межфолликулярное пространство невозможно, так как стенка фолликула лишена каких-либо пор или отверстий. Более того, если тиреоглобулин попадает в кровь в нерасщепленном состоянии, он может оказаться антигеном и вызывает процесс аутоиммунизации. Поэтому фаза выведения тиреоглобулина начинается расщеплением тиреоглобулина под влиянием фермента типа катепсинов. Тиреоглобулин еще находясь в полости фолликула подвергается протеолизу. Продукты протеолиза, в том числе йодированные аминокислоты, реадсорбируются клетками фолликулярного эпителия и здесь расщепляются лизосомальными ферментами. Процессы продукции гормонов и их отдача в ток крови совершается одновременно. Следовательно, чередования фаз продукции и выделения не происходит.
Транспорт тиреоидных гормонов
Тиреоидные гормоны (тироксин Т4 и трийодтиронин ТЗ) циркулируют в крови в связанном с белком виде — тироксинсвязывающим глобулином и лишь в очень небольших количествах — в свободном виде, и только свободные ТЗ и Т4 являются биологически активными. Секретируемые в щитовидной железе Т3 и Т4 имеют неодинаковую активность: Т3 в 4-5 раз более активен, чем Т4. Поэтому в тканях Т4 переходит в Т3 и биологическое действие тиреоидных гормонов более чем на 90-92% осуществляется за счет Т3. Механизм действия тиреоидных гормонов Тиреоидные гормоны (тироксин T4 и трийодтиронин - Т3), так же как и стероидные, легко диффундируют через липидную клеточную мембрану и взаимодействуют с внутриклеточными рецепторами. По другим данным, сначала они взаимодействуют с рецептором на плазматической мембране и лишь после этого попадают в цитоплазму, где комплексируются с белками, образуя так называемый внутриклеточный пул тиреоидных гормонов. Сродство внутриклеточных рецепторов к трийодтиронину (Т3) в 10-20 раз больше, чем к Т4 (тироксину). Поэтому биологическое действие в основном осуществляется Тз, а Т4 превращается в Тз, который связывается с ядерными, цитоплазматическим и митохондриальными рецепторами. Тиреоидные гормоны регулируют синтез белка на уровне транскрипции, и это их действие, обнаруживаемое через 12-24 ч, может быть блокировано введением ингибиторов синтеза РНК. Помимо внутриклеточного действия, тиреоидные гормоны стимулируют транспорт глюкозы и аминокислот через клеточную мембрану, непосредственно влияя на активность некоторых локализованных в ней ферментов. Таким образом, специфическое действие гормона проявляется лишь после комплексирования его с соответствующим рецептором. Это вызывает последовательную цепь пострецепторных взаимодействий, заканчивающихся проявлением специфического биологического эффекта гормона.
Физиологическое действие йодсодержащих гормонов щитовидной железы Йодсодержащие гормоны щитовидной железы резко усиливают окислительно-восстановительные и протеолитические процессы. Под их влиянием усиливается основной обмен (при введении человеку 1 мг тироксина суточный расход энергии повышается примерно на 1000 ккал). Основным обменом называется минимальный расход энергии в единицу времени, достаточный для поддержания жизни (поддержание температуры тела, обеспечения энергией сердечных сокращений, дыхательных движений и т.д.). Измерение интенсивности основного обмена производят натощак, в покое. В этих условиях почти все энергетические изменения в организме сводятся к образованию тепла, так что общий обмен можно описать как окисление энергетических запасов с выделением тепла. Интенсивность основного обмена можно выражают числом калорий на 1 м2 поверхности тела в 1 час. Среднее значение интенсивности основного обмена у взрослого мужчины равно 40 кал/м2/час, что составляет 1700 кал в сутки. Введение тиреоидных гормонов сопровождается повышением интенсивности основного обмена, что сопровождается усилением потребления О2 и образованием тепла. Таким образом, реализуется калорический эффект тиреоидных гормонов. Интенсификация основного обмена проявляется разнообразно – развивается тахикардия, расширяются сосуды кожи, усиливается потоотделение, повышается возбудимость ЦНС, ЖКТ. Следствием интенсификации окислительно-восстановительных процессов является возрастание потребления О2 и выделения СО2. В результате этого организм становится чувствительным к недостатку кислорода; при этом плохо переносится пребывание на больших высотах. Тироксин усиливает расходование всех видов питательных веществ – углеводов, жиров, белков. Под его влиянием возникает интенсивное потребление тканями глюкозы крови. Этот процесс компенсируется усилением распада гликогена в печени и мышцах. Интенсификация липолиза приводит к тому, что в мышцах сохраняется только1/3, а в печени 1/2 нормального количества жира. Усиленный распад белков ведет к увеличению количества азота в моче. Таким образом, йодсодержащие гормоны щитовидной железы оказывают выраженное метаболическое действие, участвуют в процессах термогенеза. У здоровых людей ЩЖ принимает участие также в контроле за весом тела. При повышенном потреблении пищи активность ее увеличивается, образование Т3 усиливается, что приводит к повышению скорости метаболизма в организме. Наоборот, при недоедании активность ЩЖ снижается, приводя к замедлению метаболизма. Тиреоидные гормоны необходимы для нормального умственного и физического развития. Наряду с гормоном роста, вырабатываемым в гипофизе, они отвечают за нормальное развитие костей скелета. Недостаток гормонов ЩЖ в детском возрасте приводит к прекращению роста, а дефицит их при беременности - к недоразвитию мозга будущего ребенка – развитию кретинизма. Йодсодержащие гормоны оказывают стимулирующее действие на ЦНС, способствуют нормальному функционированию симпатической нервной системы, особенно ее адреналовому компоненту, благодаря стимулированию синтеза β-адренорецепторов в миокарде, скелетных мышцах, жировой ткани и т.д. Физиологические эффекты тиреоидных гормонов изучаются и при анализе их гипо- и гиперсекреции, эффекты которых представлены в табл. 7.1.
Таблица 7.1 Дата добавления: 2015-02-06 | Просмотры: 2228 | Нарушение авторских прав |