|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология
|
Время свертывания крови у разных видов рыб, с
| Вид рыб
| Время свертывания крови, с
| Условия
| | Карп, лещ, густера
| 600-840
| Выловлены из природных водоемов
| | Плотва, язь
| 300-380
| То же
| | Ерш, окунь, судак
| 120-180
| То же
| | Радужная форель
| 150-250
| Выловлена из искусственных бассейнов
|
Таким образом, организм рыб надежно защищен от больших кровопотерь. Зависимость времени свертывания крови рыб от состояния нервной системы является дополнительным защитным фактором, поскольку крупные кровопотери возможны скорее всего в стрессовых ситуациях (нападение хищника, драки).
§28. КРОВЕТВОРЕНИЕ
Постоянное присутствие в крови незрелых форм эритроцитов и белых клеток свидетельствует о том, что гемопоэз у рыб протекает очень интенсивно. Кроветворение у рыб специфично не только по интенсивности, но и по локализации процесса. У рыб отсутствует красный костный мозг - основной орган кроветворения высших позвоночных. У рыб функцию гемопоэза выполняют другие органы. К ним относятся почки, сердце, жабры, селезенка, образования лимфоидной ткани.
Почки у рыб выполняют не только выделительную, как у других позвоночных, но и кроветворную функцию. Например, у сеголетков карпа в гистоструктуре почек 60 % площади приходится на ретикулярный синцитий и только 40 % - на выделительные канальцы. Наиболее активный гемопоэз у карповых рыб, некоторых окуней и сомов происходит в периферической части почек. У большинства рыб повышенная кроветворная активность зафиксирована в каудальной и головной частях почки.
Селезенка рыб обеспечивает образование клеток эритроидного ряда. Поэтому селезенка функционально напоминает красный костный мозг высших позвоночных. Многие исследователи считают селезенку и органом лимфогранулопоэза.
Сердце рыб внутри выстлано однослойным плоским эпителием, который также причастен к процессу кроветворения, как и эндотелий кровеносных сосудов рыб.
Наличие лимфоидных образований в слизистой пищеварительной трубки рыб позволяет ассоциировать с кроветворением и желудочно-кишечный тракт. Однако многие исследователи считают, что его лимфоидные образования являются резервуарами белых клеток, за счет которых происходит постоянное пополнение пула белой крови, циркулирующей по сосудам.
Важное место в процессе образования белой крови отводится тимусу и лимфоидному органу.
Тимус имеет различное расположение и морфологию у разных видов рыб. Так, например, у скатов он хорошо оформлен и расположен позади брызгальца. У щуки и угря тимус представлен уплотнением медиальной стенки жаберной полости, В отличие от млекопитающих тимус у рыб функционирует всю жизнь. Он имеет корковый и мозговой слои. Мозговой слой отвечает за образование Т-лимфоцитов. Эндокринная функция тимуса рыб не изучена.
Лимфоидный орган представляет собой скопление лимфоидной ткани и располагается у костистых рыб за черепом, перед почками. Элементы лимфоидного органа заполняют пространства между позвонками и укрыты соединительной тканью.
У осетровых рыб лимфоидный орган находится под крышей черепа (рис. 6.5).
Он представляет собой парное, почти округлое образование, У трехлетней белуги его масса достигает 0,8 % массы тела. На гистосрезах лимфоидного органа всегда присутствуют нейтрофилы, эозинофилы, базофилы, тромбоциты и даже незрелые эритроциты, т. е. лимфоидный орган причастен к процессу формирования всех клеток крови.
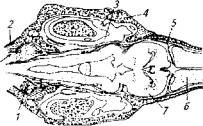
Рис. 6.5. Топография лимфоидного органа у осетровых:
1 - лимфоидный орган; 2- черепная коробка; 3-полукружные каналы; 4-хрящ; 5-мозговая оболочка; 6-передний мозг; 7- средний мозг
Ретикулярный синцитий обнаружен во всех висцеральных органах, за исключением печени.
Таким образом, кроветворная функция характерна для многих органов рыб. Интенсивность кроветворения разных органов характеризует табл. 6.14.
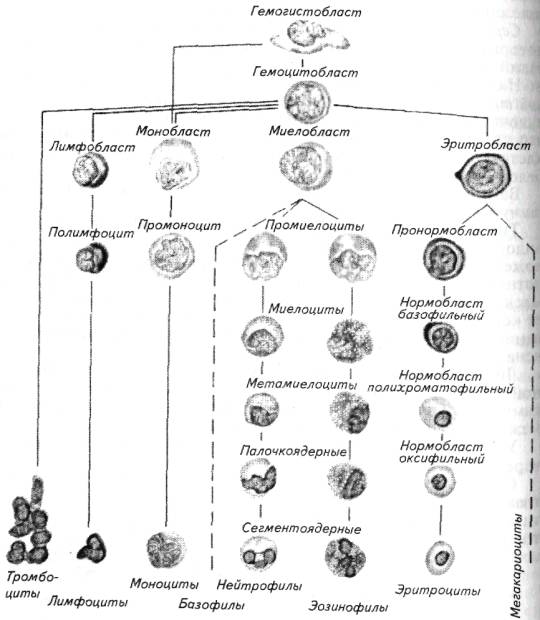
Рис. 6.6. Происхождение отдельных клеток крови рыб
6.14. Содержание форменных элементов в различных органах кроветворения и сосудистой крови, %
| Клетки
| Жабры
| Почки
| Желудочно-кишечный тракт
| Сердце
| Селезенка
| Лимфоидный орган
| Сосудистая кровь
| | Эритробласты
| 7,6
| 3,3
| -
| 2,5
| 2,5
| 3,3
| 1,0
| | Эозинофилы
| 2,0
| -
| -
| -
| 5,0
| -
| 2,0
| | Эозинофилы
| 3,5
| 6,0
| 1,0
| 8,0
| 15,2
| 18,2
| 1,8
| | Гемоцитобласты
| 1,0
| 2,9
| -
| -
| 3,0
| 2,5
| 1,5
| | Миелобласты
| 1,0
| 2,0
| -
| -
| 2,0
| -
| 1,0
| | Промиелобласты
| 2,5
| 2,2
| -
| 1,0
| 2,5
| 1,0
| 3,5
| | Миелоциты
| 3,6
| 3,4
| -
| 2,5
| 5,2
| 7,0
| 2,7
| | Метамиелоциты
| 3,0
| 4,8
| 2,0
| 2,11
| 5,2
| 3,5
| 4,0
| | n-Гранулоциты
| 6,5
| 9,6
| 2,0
| 5,0
| 9,1
| 20,0
| 10,6
| | С-Гранулоциты
| 4,0
| 11,6
| 1,0
| 12,2
| 9,9
| 3,5
| 12,2
| | Лимфоциты
| 61,3
| 53,6
| 93,5
| 38,4
| 65,5
| 39,0
| 54,7
| | Моноциты
| 4,0
| -
| 0,5
| 1,7
| 2,0
| 2,0
| 5,0
|
Жабры как хрящевых, так и костистых рыб содержат клетки ретикулярного синцития, в которых всегда присутствуют все типы клеток крови. Поэтому жаберный аппарат за счет синцития, а также эндотелия кровеносной системы жабр участвует в кроветворении у рыб.
Все клетки крови имеют единое происхождение от гемоцитобласта (рис. 6.6). Созревание отдельных форм лейкоцитов протекает в лимфоидной ткани пищеварительной трубки, лимфоидном органе, тимусе. Созревание тромбоцитов, скорее всего, происходит в селезенке.
Интенсивность гемопоэза зависит от внешних и внутренних факторов. Так, при дефиците кислорода наблюдается усиление эритропоэза; пониженная температура воды и ее загрязненность органическими веществами стимулируют лейкопоэз за счет фагоцитирующих форм. Инфицирование рыбы, кроме фагоцитоза, усиливает и морфогенез лимфоцитов.
§29. ФУНКЦИИ ОТДЕЛЬНЫХ КЛЕТОК КРОВИ
В литературе можно найти обширный материал по количественной характеристике клеток крови рыб. Однако большинство авторов очень сдержанны в отношении функциональных характеристик клеток крови рыб. Не всегда понятно происхождение отдельных клеток крови рыб и их функциональное предназначение. Поэтому представляется полезным сделать специальное обобщение по данному разделу физиологии рыб.
Эритроциты рыб (рис. 6.7) подобно эритроцитам высших позвоночных связывают с функцией переноса кислорода, поддержанием кислотно-щелочного равновесия, в меньшей мере с транс сортированием диоксида углерода. Помимо этого эритроциты рыб причастны к транспортированию низкомолекулярных органических соединений и механизму дезинтоксикации организма.
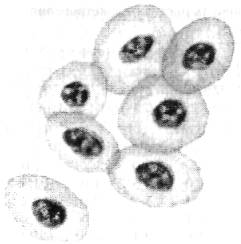
Рис. 6.7. Эритроциты рыб
Молодые формы эритроцитов рыб способны к фагоцитозу. Практически у всех видов рыб эритроциты имеют овальную (или близкую к ней) форму с продольным размером от 8 мкм у щуки до 18 мкм у карася.
Лейкоциты крови рыб представляют собой менее однородную по сравнению с эритроцитами группу клеток с большим разнообразием линейных размеров (4-20 мкм), различной структурой ядра, цитоплазмы и даже клеточной оболочки. Все белые клетки классифицируют на основе их способности при окрашивании демонстрировать гранулярную или негранулярную структуру цитоплазмы (рис. 6.8).
Не всегда ясно происхождение отдельных форм лейкоцитов. Отсюда и естественная осторожность при оценке их функционального назначения.
Лимфоциты являются самой представительной у рыб группой и занимают до 99 % лейкоцитарного ряда. Они делятся на мелкие (4-8 мкм) и крупные (9-12 мкм) формы. Лимфоциты имеют очень крупное ядро, занимающее большую часть объема клетки. Они постоянно присутствуют в лимфе. Лейкоциты легко мигрируют из кровеносной системы в лимфатическую систему и обратно. Возможно, этим объясняется неоднородность и противоречивость экспериментальных данных о количестве лимфоцитов в кровяном русле.
Несмотря на то что лимфоциты обнаружены в мазках почек, селезенки, жабр, тем не менее их происхождение связывают с тимусом. Клетки, производящие антитела, чаше обнаруживают в мазках передней почки. У рыб лимфоциты функционально можно отнести к двум группам. В-клетки имеют рецепторы к гамма-глобулинам (аналог В-лимфоцитов высших позвоночных). Т-группа включает, как минимум, два вида клеток - киллеров и их хелперов. Супрессоры и клетки памяти крови рыб не описаны в литературе, но без их наличия трудно объяснить реакции иммунитета у рыб. Иммунокомпетентность лимфоцитов подчеркивает продолжительность их жизни. Для Т-клеток она составляет 90 дней, для В-клеток - 4-5 дней.
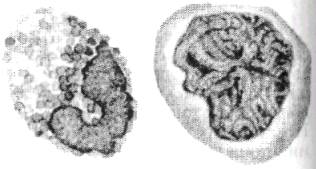
Рис. 6.8. Пример белых клеток крови с гранулированной (гранулоцнт) и негранулированной (агранулоцит) структурой
Лимфоциты рыб обеспечивают и специфические иммунологические реакции, реакции отторжения. Кроме того, лимфоциты являются предшественниками макрофагов, фибробластов, "мачтовых" клеток.
Лимфоциты рыб не способны к фагоцитозу. Все литературные указания на этот счет не более чем ошибка. При использовании несовершенной техники идентификации лимфоцитов их легко спутать с моноцитами и тромбоцитами и ошибочно приписать несвойственные им функции.
Моноциты составляют 0,1 % общего количества крови. По достижении стадии зрелости через 48 ч они покидают циркуляционное русло и превращаются в макрофаги тканей. По этой причине ряд исследователей отказываются признавать моноциты самостоятельной группой и считают их "гематогенными" макрофагами у костистых рыб. Однако экспериментально установлена выраженная способность этих клеток к хемотаксису по отношению ко многим веществам и корпускулярным структурам, а также их высокая фагоцитирующая активность.
Нейтрофилы - довольно постоянная группа клеток у рыб. Например, у карповых рыб их количество составляет 150 тыс/мм3, или 25 % всех лейкоцитов крови. Это крупные (10-12 мкм) клетки с круглым или овальным ядром полиморфной структуры (отсюда название - "полиморфноядерные" клетки). У рыб нейтрофилы - основные фагоцитирующие клетки, быстро реагирующие на очаг воспаления, инфицированный микроорганизмами. Утверждение о хемотаксисе нейтрофилов, по крайней мере у костистых, спорно. Хорошо изучена нейтрофилия рыб при стрессах.
Базофш1ы обнаружены в крови не всех рыб. На это влияют как внешние, так и внутренние факторы. В тех случаях, когда базофильные клетки удается обнаружить, их размеры оцениваются 10-15 мкм. Количество базофилов составляет от 1 % у австралийской двоякодышащей рыбы до 9 % у карася и карпа. Естественно, функции базофилов изучены недостаточно. Гистохимические методы исследований позволяют говорить об их причастности к синтезу гистамина и гепарина.
Эозинофилы в циркуляторном русле обнаруживаются редко, но в кроветворных органах присутствуют постоянно. У некоторых морских рыб они составляют до 40 % всех белых клеток крови. Есть сообщения о том, что в крови золотой рыбки они занимают 8 %. Это крупные клетки (10-12 мкм) с палочко- или сегментоядерным содержимым. У карася, золотой рыбки и гуппи в экспериментальных условиях эозинофилы фагоцитировали бактериальные клетки. Замечены скопление эозинофилов в очаге воспаления и рост их общего количества до 20% при стрессе. Однако не исключено, что в этих случаях эозинофилы путали с нейтрофилами. Что не подлежит сомнению, так это наличие тканевых эозинофилов, которые в большом количестве обнаружены в жаберном эпителии, коже и в слизистой желудочно-кишечного тракта.
Макрофаги у рыб обнаружены в различных органах, покровных тканях, но не в крови. Клетки отличаются большими размерами (20-40 мкм). В то же время они очень подвижны и не имеют постоянной формы. Цитоплазма макрофагов образует псевдоподии, как у амебы. В результате клетки могут приобретать вытянутую форму с малым размером в поперечнике. Последнее позволяет макрофагам свободно мигрировать по всему организму.
Доказана высокая фагоцитирующая активность макрофагов в почке, селезенке, сердечной мышце, но не в тимусе. Причем отмечена их способность как к фагоцитозу, так и к пиноцитозу. В цитоплазме макрофагов обнаруживают остатки бактериальных клеток, эритроцитов, а также мелких паразитов. Макрофаги рыб активны и в отношении некротизированной ткани и связаны с выработкой гамма-глобулинов. Макрофаги обладают хемотаксисом, поэтому могут элиминировать различные инородные химические вещества.
Происхождение макрофагов рыб не изучено. Тем не менее их образование ассоциируют с ретикулярным синцитием кроветворных органов.
Плазматические клетки рыб, скорее всего, развиваются, как и у млекопитающих, из В-лимфоцитов. Это крупные розоподобные клетки, цитоплазма которых содержит гамма-глобулины. Красно-фиолетовое ядро клеток имеет облаковидную или колесовидную структуру хроматина. Цитоплазма располагается широким кольцом вокруг ядра и окрашивается в густой синий цвет. Функции плазматических клеток рыб не изучены. Однако наличие глобулина в цитоплазме не оставляет сомнений в том, что они причастны к реакциям специфического иммунного ответа.
"Мачтовые" клетки являются производными лимфоцитов и отличаются базофильной цитоплазмой. Считается, что эти клетки причастны к развитию анафилактического шока, хотя безоговорочно это явление у рыб исследователями не принимается, Тем не менее в экспериментах на костистых удалось получить после сенсибилизации явление, похожее на анафилаксию на введение в организм рыбы лошадиной сыворотки и яичного альбумина. В протоплазме "мачтовых" клеток обнаруживается гистамин.
"Шнуровидные" клетки -это эозинофильные клетки, обнаруженные на поверхности жаберного эпителия и в слизистой кишечника костистых рыб. Они имеют вытянутую перпендикулярно поверхности ориентированную форму с гранулированной в виде тяжей - "шнурков" - цитоплазмой. "Шнуровидная" клетка посредством поры соединяется с эпителиальными клетками. Клетки имеют тонкую (1 мкм) фиброзную капсулу. Предполагается, что "шнуровидные" клетки происходят от "бродячих" лейкоцитов, физиологическая роль этих клеток не изучена.
функции тромбоцитов рыб изучены недостаточно. Однако имеющейся в специальной литературе информации достаточно, чтобы не ограничиваться механическим переносом на рыб функций, известных у млекопитающих. Принято считать, что у высших позвоночных эти клетки обеспечивают первый этап свертывания крови, т. е. образование тромбопластина. У рыб первый этап свертывания крови более зависим от тканевой тромбокиназы, которая в достаточных количествах выделяется стенками кровеносных сосудов при их повреждении. Более того, кожная слизь рыб имеет высокую тромбокиназную активность.
Таким образом, роль тромбоцитов у рыб в триггерном механизме свертывания крови выглядит скромной. Это подтверждается и исключительно высокой скоростью свертывания крови при ее контакте со слизью кожи. Кроме тромбокиназы кожная слизь содержит кинины - полипептиды, которые активируют фактор Хагемена (фактор XII) и фактор XI системы коагуляции.
Функции тромбоцитов рыб не ограничиваются участием в первом этапе свертывания крови. Установлено, что при формировании тромба на третьем этапе свертывания крови цитоплазма тромбоцитов шнуруется с образованием сетчатой структуры, которая улавливает клетки крови.
Внутривенное введение мелкодисперсного углерода доказывает причастность тромбоцитов и к фагоцитозу. Частицы углерода хорошо просматриваются в цитоплазме не только классических фагоцитов, но и тромбоцитов рыбы.
Дата добавления: 2014-12-11 | Просмотры: 1811 | Нарушение авторских прав
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 34 | 35 |
|