|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология
|
Вторичные половые признаки, имеющие большое значение при распознавании пола партнера
| Вид рыб
| Признаки, принимаемые во внимание
| | самцом при распознавании самки
| самкой при распознавании самца
| | Хромис красавец
| Поза покорности самки. Следование самки за самцом
| Яркая окраска. Ритуальное ухаживание самца (преследование).
| | Петушки
| Окраска самки: металлический отлив, появ-ление полос на теле. Надувание жаберного аппарата. Следование за самцом
| Яркая окраска тела, большие расправленные плавники. Специфическое ухаживание сам-ца. Ритуальные удары хвостом
| | Гуппи
| Более крупное тело самки. Большое брюшко. Отсутствие агрессивного поведения
| Меньшие размеры, хорошо развитые плавники. Преследование самки и кружение вокруг нее
| | Колюшка
| Отсутствие красного пятна на выпуклом брюшке. Ритуальное поведение самки: плавает наклонно вниз головой, вибрируя телом
| Красное брюшко. Зигзагообразные движения тела. Ложные заходы в гнездо для нереста
| | Горчак
| Наличие яйцеклада
| Окраска самца
| | Меченосец
| Выпуклое брюшко, отсутствие меча. Отсутствие агрессивного поведения
| Наличие меча и копулятивного органа. Ритуальные приставания
|
Вторичные половые признаки хорошо выражены у пелагических рыб. Рыбы донные, рыбы, обитающие в мутных водах, пещерах. т. е. рыбы с плохим зрением, как правило, имеют плохо выраженные вторичные половые признаки. У этих рыб решающее значение при распознавании половых партнеров приобретают феромоны, а также звуковая, тактильная и электрическая сигнализация. Узнаваемый, индивидуальный портрет создают и гидродинамические поля, сопровождающие движение каждой особи в воде (Рис. 15.11).
У рыб известны примеры группировок парных рыб. Рыбы образуют пары для нереста и последующего ухода за молодью. Для этих рыб характерны сложные ритуальные нерестовые танцы. Некоторые из них строят гнездо ухаживают и охраняют молодь (например, судак, колюшка многие лабиринтовые рыбы)' Чаше всего такие рыбы вне сезона размножения состоят членами больших стай (лососевые сомовые, осетровые).
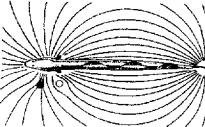
Рис. 15.11. Гидродинамический портрет рыбы
В редких случаях рыбы образуют парные ассоциации, имеющие и другой биологический смысл. Например, между муреной и губаном складываются симбиотические отношения (рис. 15.12).
Губан тщательно обследует кожу и даже пасть мурены. При этом он пожирает паразитов, выкусывает кусочки омертвевшей ткани, очищает ротовую полость от гниющих остатков корма. Губан не одиночка среди рыб. Описано, по крайней мере, два десятка видов рыб, выполняющих санитарную функцию.
Другим проявлением парных ассоциаций рыб является комменсализм (нахлебничество).
Типичным примером комменсализма служит ассоциация акул и рыб-лоцманов. Рыбы-лоцманы сопровождают акулу круглосуточно. Они провоцируют хищника к атаке, совершая первыми броски на жертву. Рыбы-лоцманы имеют небольшие размеры тела. Поэтому они довольствуются мелкими остатками пищи, которые акулу не интересуют.
Подобным образом удовлетворяет свои пищевые потребности и рыба-прилипала, которая отделяется от акулы только для того, чтобы подобрать объедки.
На рис. 15.13 показан пример парных ассоциаций рыб, название которым сформулировать довольно сложно.

Рис. 15.12. Пример симбиотических отношений между муреной и чистильщиком губаном
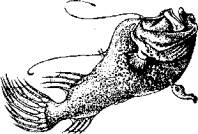
Рис. 15.13. Самка и приросший к ее телу самец (глубоководный удильщик)
У глубоководного удильщика очень большая разница в размерах тела между самкой и самцом. Крошечный самец прирастает к телу самки. Большую часть жизни самец выступает как паразит, так как питается соками тела самки. Однако в период размножения он исправно выполняет свои прямые обязанности, находясь в столь необычном положении. Эта парная ассоциация рыб может быть определена как сексуальный паразитизм.
Среди рыб распространен и истинный паразитизм. Этот вид взаимоотношений с другими представителями водных животных отличает прежде всего миксин, миног и многочисленных представителей сомовых рыб. Мелкие сомики из семейства Trichomycteridae - представители ихтиофауны Южной Америки - прогрызают кожу крупных рыб и питаются их кровью.
Сом-стегофил паразитирует на жабрах крупных рыб при помощи специальных присосок. Другой сомик - ванделия - представляет опасность даже для людей. Обычно он паразитирует в мочеточниках крупных рыб, но может проникать в мочеполовую систему и человека.
Очевидно, что в парных ассоциациях рыб далеко не всегда оба партнера имеют какую-то выгоду. Однако даже в самом благоприятном случае она касается всего лишь пары представителей вида. Многочисленные ассоциации оказывают более выраженное влияние на состояние популяций.
§58. СТАЙНЫЙ ОБРАЗ ЖИЗНИ
Крупные группировки рыб облегчают выполнение жизненных задач индивидууму (защиту от опасности, поиск пиши, ориентацию при миграциях). Стая - это группировка близких по возрасту и физиологическому состоянию особей рыб одного вида, объединяющихся на достаточно продолжительный отрезок времени. Стайными являются в основном пелагические виды.
Вероятно, главная биологическая задача стаи - самосохранение. В стае рыбы становятся более пугливыми. Стая проявляет высокую настороженность и активно избегает раздражителей, к которым вне стаи особи относятся терпимо. К стае хищнику подобраться сложнее, чем к одиночной рыбе. Отдельные движущиеся Рыбы в стае трудноразличимы для хищника, так как он не может остановить взгляд на одной особи. Бросок хищника на стаю, как правило, не приносит результата, поскольку стая делится на две части и перед хищником оказывается пустота. Опытный хищник может длительное время кружить вокруг стаи, не нападая на нее. Он стремится отбить отдельную рыбу от стаи и лишь после этого производит результативный бросок.
Стая рыб раньше замечает опасность. Стаю настораживает все и необычное: крупный объект в воде и на берегу, тень от крупного объекта на берегу, посторонние звуки, гидродинамические удары, электромагнитные поля, изменение рН и химического состава воды. Стая рыб менее склонна к исследовательскому поведению, заменяя таковое активным избеганием всего нового.
Стайный образ жизни обеспечивает лучшую выживаемость как молоди, так и взрослых рыб. Стая эффективнее в поиске корма, распознавании опасностей и нахождении миграционных путей.
Количество особей в стае может быть самым различным: от нескольких десятков (сазан, треска) до тысяч (сельди) и сотен тысяч (анчоус) рыб.
В зависимости от экологической ситуации и биологической пели структура стаи и ее форма могут сильно видоизменяться (рис. 15.14).
Как правило, у большинства видов рыб при быстром движении стая имеет клиновидную форму, при питании - округлую. Реакция на опасность сильно различается у разных видов рыб. Например, стая анчоуса в случае опасности превращается в плотный шар, а стая скумбрии опускается на глубину и там рассеивается.
Для морских рыб обычны скопления стай. Скопления рыб по биологическому значению определяются как нерестовые, нагульные, зимовальные и миграционные.
Нерестовые скопления возникают в местах размножения рыб. Они достаточно однородны и представлены только половозрелыми особями.
Нагульные скопления рыб формируются в кормных местах, образуются из разновозрастных особей с большим удельным весом молоди.
Миграционные скопления еще менее однородны, так как возникают на путях движения рыб к нерестилищам, местам зимовки или местам откорма. Этот тип скоплений может включать в себя даже стаи разных видов рыб.

Рис. 15.14. Организационные типы стаи рыб:
1-ходовая; 2 -стая кругового обзора; 3 и 3а - оборонительная стая; 4-стая планктонофагов на питании; 5 и 5а - стая хищных рыб при питании
Рис. 15.15. Скопление хамсы в поисках Керченского пролива:
а, 6, в, г - четыре фазы амебоподобной деформации стаи
Скопления объединяют миллионы особей в единый организм (рис. 15.15). Так, мурманская сельдь, черноморская ставрида, сардинелла, тихоокеанские лососи образуют скопления, которые не поддаются подсчету без специальных инструментов.
В отдельные годы черноморская хамса образует скопления, занимающие площадь в сотни тысяч квадратных метров при высоте до 20 м. Протяженность косяков черноморской кефали достигает 100км.
Такие скопления рыбы регистрируют при помощи авиации или из космоса. Их образование связывают с миграциями, нагулом, нерестом и зимовкой.
Скопления могут быть образованы стаями рыб разных видов. При этом взаимоотношения между стаями не носят хаотичного характера. Между стаями осуществляется обмен информацией по нескольким каналам, что подтверждает организованный характер реагирования скоплений на внезапное появление опасности (крупные хищники, морские суда). Даже простые дыхательные движения рыб сопровождаются особыми сигналами (рис. 15.16).
Стая имеет свое индивидуальное электрическое, а также гидродинамическое поле. Она оставляет после себя химический след.
Результаты наблюдений показали, что периодические контакты имеют определенный биологический смысл. Если по какой-либо причине стая или косяк рыб не встречает на пути своей
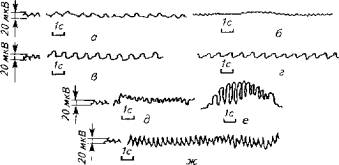
Рис. 15.16. Низкочастотные электрические поля, сопровождающие дыхание рыб: а -вьюн; б-красноперка; в - ратан; г -линь; д- карась; е- цихлозома; эк - вуалехвост (В. Р. Протасов, 1978)
миграции другие стаи, то стая (косяк) начинает избегать это место, прокладывая другие маршруты движения в районы, где возможен контакт с другими стаями. Новый маршрут движения будет закреплен, если контакты с другими стаями на новом маршруте постоянны и приходятся на определенное время.
Поведение стаи как единого целого плохо поддается пониманию. Так, скопление рыб, занимающее площадь в несколько гектаров (например, черноморская хамса), движется как один организм. Движения головных стай синхронизированы с движениями стай, замыкающих скопление. Движение многотысячного скопления рыб напоминает движение амебы (см. рис. 15.15).
В стаях между отдельными членами складываются взаимоотношения двух типов: равноправные (стая не структурирована, например у хамсы, верховки) и ранжированные (с вожаком, например тунцы, окуневые, кефаль). Стая может быть привязана к определенному месту водоема (территориальная стая). Другой тип стаи (ходовая стая) постоянно перемещается по водоему. В основе поддержания целостности стаи лежит реакция следования, т.е. врожденная реакция отдельной особи следовать за другими. Как структурированная, так и неструктурированная стая имеет две зоны: внутреннюю и наружную.
На факторы внешней среды реагирует ядерная часть стаи. Рыбы, находящиеся на периферии стаи, подражают ядру, т. е. следуют за ним. Эксперименты с моделями показали, что стая как единое целое начинает движение в том случае, если перемещение инициировали не менее 30 % особей ядра. Движению стаи предшествует своеобразное возмущение ядра. И лишь когда моторная активность ядра достигнет "критической массы", стая начинает движение.
Чаще всего движение стаи носит лавинообразный характер.
Обнаружив корм или источник опасности, стая совершает резкий бросок, который затухает после поедания корма или уклонения от опасности. Однако и в отсутствие стимула стая движется волнообразно, так как даже исследовательское поведение отдельных членов стаи привлекает внимание других членов.
Примитивисты полагают, что поведением животных управляют безусловные и условные рефлексы. Их правота заключена лишь в одном: действительно поведение животных складывается в результате непростого суммирования генетически детерминированных локомоторных реакций и новых форм поведения, возникших в результате личного восприятия факторов среды обитания.
Мы исходим из того, что рефлексы, вне всякого сомнения, играют важную роль в поведении и функционировании животного организма в целом.
Однако необходимо признать, что одними рефлексами невозможно объяснить поведение даже беспозвоночных животных. Поведение животных развивается как в филогенезе, так и в онтогенезе. Со временем животные учатся, приобретают опыт, становятся способными принимать неожиданные решения и выполнять очень сложные действия, понимание которых невозможно с позиций концепции "стимул - ответная реакция".
§59. ОПЫТ, НАУЧЕНИЕ, РАЗВИТИЕ ПОВЕДЕНИЯ В ОНТОГЕНЕЗЕ
Опыт. В процессе онтогенеза экспрессия генов и синтетические процессы, лежащие в основе деления клеток и органогенеза, находятся под влиянием окружающей среды. В период роста молоди рыб многие гены сохраняют свою высокую активность и многие процессы растущего организма испытывают прессинг как со стороны самого организма, так и извне. Это относится и к эффекторным структурам двигательных реакций.
Экспериментальные данные свидетельствуют о том, что среда влияет как на развитие, так и на проявление поведенческих реакций рыб. Врожденные рефлексы и инстинкты корректируются. Отсюда следует, что не существует поведенческих реакций, независимых от генетики рыбы, как не существует поведения, независимого от окружающей рыбу среды. Другим следствием является то, что любое взаимодействие индивидуума с окружающей средой предопределяет потенциальную возможность изменчивости этого индивидуума.
Влияние среды на поведение индивидуума реализуется как через раздражение специализированных клеток (рецепторный аппарат), так и путем прямого воздействия на прочие клетки организма, включая клетки центральной нервной системы. Например, изменение концентрации кислорода в воде приводит к изменению поведения рыбы. При низкой концентрации кислорода рыба прекращает питаться, уходит в другое место водоема, поднимается к поверхности и начинает глотать атмосферный воздух.
Перечисленные поведенческие акты происходят под влиянием окружающей среды, изменения которой не рецепцируются специальными сенсорными клетками. Тем не менее в данном случае рыба получает информацию о заморном участке водоема и о возможных последствиях пребывания в нем.
В другом участке водоема рыба (например, карась) может при помощи сенсорных органов (органов зрения, обоняния или акустико-латеральной системы) обнаружить хищника и уклониться от прямого контакта с ним. В обоих случаях карась приобретает опыт. Некоторый опыт приобретается как результат изменения гормонального статуса рыбы.
Таким образом, рыбы приобретают опыт не только на основе сенсорной информации из внешней среды. Но в любом случае опыт - это изменение функциональной активности мозга (а возможно, и его морфологической структуры), возникшее под влиянием информации из внешней или внутренней среды организма. Некоторый опыт сохраняется в течение короткого времени, а какой-то опыт остается на всю жизнь. Опыт отражает коррекцию филогенетических стереотипов поведения на основе онтогенетической биологической целесообразности. Чем выше эта целесообразность, тем вероятнее генетическое закрепление опыта как этологического приобретения.
Научение. Оно имеет прямое отношение к приобретению опыта. Существует ошибочное мнение, что данное нейрофизиологическое явление характерно только для высших позвоночных, имеющих большие полушария и кору. На самом деле это неверное представление. В нашем понимании научение -это изменения в мозге (функциональные и, возможно, морфологические) под влиянием информации извне, приводящие к достаточно длительно продолжающимся коррекциям поведения, сформированного на основе индивидуального опыта.
Научение относится ко всем видам поведения животных. Существенной особенностью научения является то, что его последствия могут проявляться в любое время: сразу в ответ на действие фактора окружающей среды, или с некоторым запозданием, или не проявиться вовсе на фоне конкретного физиологического состояния организма. Поведенческая реакция может быть цикличной и повторяться многократно в отсутствие стимула. Данное явление теория рефлексов объяснить не может.
Очевидно, что многие факторы среды вообще не приводят к изменениям поведения рыб в будущем. Одни события остаются не замеченными рыбой, другие не достигают центров, отвечающих за принятие решения вследствие фильтрации афферентного потока. Очевидно и то, что определенные раздражители выступают в роли пусковых механизмов проявления поведенческих актов различной сложности. Остается загадкой, как отличают рыбы пусковые (сигнальные) раздражители от несущественных событий.
Предполагают, что в основе научения животных дифференцировать раздражители на значимые в данный момент, менее значимые и несущественные лежат два явления.
Во-первых, генетическая предрасположенность к определенным раздражителям, ассоциируемым с конкретными действиями животного.
Во-вторых, изменчивость реактивности нервной системы животного после многократных повторных реакций на один и тот же раздражитель.
Предрасположенность к научению проявляют практически все позвоночные и многие беспозвоночные (примером служат пчелы) животные. Особенно наглядно она проявляется в опытах с отвергаемыми (негативными) стимулами, опытах, ставших классическими со времен Конрада Лоренца и Нико Тинбергена. Так, реакция избегания стимулов, вызывающих болевые реакции у животного, формируется значительно быстрее, чем двигательные реакции на положительные стимулы (пищевые). Более того, пищевые стимулы неодинаково эффективны при запуске локомоторных реакций, имеющих различное биологическое предназначение.
В свое время Лоренц и Тинберген предложили концепцию врожденного пускового механизма (ВПМ). Она базируется на трех постулатах:
1. Механизм ВПМ генетически детерминирован, т.е. имеет врожденный характер.
2. ВПМ выполняет роль рилизинг-фактора, запускающего (активизирующего) нервные процессы в центральной нервной системе. ВПМ может тормозить активизацию центральной нервной системы, если сигнальный раздражитель не распознан животным, а также ввиду других причин.
3. ВПМ запускает стереотипическую ответную реакцию животного на сигнальный раздражитель.
Врожденный пусковой механизм предполагает запуск стереотипической ответной реакции, отличной от рефлекса, так как она возникает на различные изменения окружающей среды, в то время как рефлекс - это ответная реакция только на вполне определенный стимул.
Классическим примером врожденного пускового механизма является реакция самца трехиглой код юшки в период нереста на красный цвет брюшка другой рыбки или модели (рис. 15.17).
Пример с колюшкой демонстрирует различия между рефлекторной реакцией рыбы на стимул и действием врожденного пускового механизма. Здесь красный цвет выступает сигнальным стимулом только в период половой активности самца, причем красная окраска должна быть нанесена на нижнюю часть модели (брюшко). В случае с рефлексом ответ должен возникать у животного независимо от физиологического состояния и местоположения красного пятна.
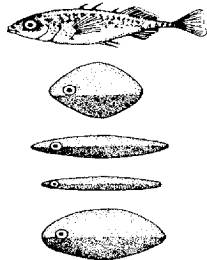
Рис. 15.17. Красный цвет брюшка (модели) в качестве сигнального раздражителя ВПМ у колюшки
Скорость научения увязывать раздражитель с действием определяется еще характером возможных последствий. Рыбы проявляют большую предрасположенность к научению в ответ на действие стимула, исходящего от хищника или другой смертельной опасности. К стимулам, не ассоциируемым с большой опасностью, развивается привыкание, т. е. реакция замедляется или даже прекращается со временем.
Предрасположенность к научению развивается в результате сочетания генетических факторов и факторов окружающей среды. У каждого индивидуума генетические и экологические факторы взаимодействуют при формировании предрасположенности к научению в общем смысле и применительно к конкретным обстоятельствам. Животные, располагающие большей предрасположенностью к научению, имеют больше шансов на выживание, дают более многочисленное потомство, которое более эффективно защищено от воздействия неблагоприятных факторов. В конечном счете, большая предрасположенность к научению закрепляется генетически, так как особи с пониженной способностью к научению чаще гибнут и не оставляют потомства.
Самой простой формой научения у животных этологи считают импринтинг. Теорию импринтинга детально разработал лауреат Нобелевской премии К. Лоренц. Он показал, что реакция следования проявля-ется у большинства позвоночных животных. Различия заключаются лишь в природе стимула и времени проявления импринтинга в раннем онтогенезе. Биологическое значение импринтинга велико. Считается, что он имеет отдаленные последствия на половое поведение животных. Импринтинг определяет способность животных распознавать сородичей, выявлять среди них близких родственников, что, в свою очередь, позволяет им избегать близкородственных спариваний и последующего вырождения из-за инбридирования.
Импринтинг у рыб не описан. Однако нет оснований для того, чтобы рассматривать его в качестве этологического проявления ароморфоза, не затронувшего класс рыб. Автор считает, что импринтинг (так хорошо изученный у птиц и млекопитающих) эволюционизировал из оптомоторной реакции рыб.
Оптомоторная реакция - это врожденное свойство животных направленно двигаться в движущемся зрительном поле. Данное явление описано не только у рыб, но и у более примитивных животных - ракообразных, насекомых.
С возникновением импринтинга у высших позвоночных оптомоторная реакция не утратилась. Она проявляется даже у человека в форме так называемого "железнодорожного нистагма". Когда человек находится в быстродвижущемся поезде, человеческий глаз непроизвольно следует за мелькающими за окном объектами.
Это же явление отмечается и у высших животных. Например, когда молодую собаку впервые берут в путешествие на автомобиле, она не в состоянии сосредоточиться на чем-то внутри автомобиля. Ее глаза рефлекторно отслеживают все движения за пределами автомобиля (с точки зрения собаки за окном движется практически все).
Из-за столь неадекватной реакции у животного возникает избыточный поток зрительной афферентации, что может привести к нервному срыву. Поэтому собаку надо постепенно приучать к поездкам на автомобиле, дозируя нагрузку.
Зрительно-двигательная реакция проявляется у животных по-разному и заключается не только в движении глаз. Отдельные особи сопровождают движения глаз поворотом головы, изгибами тела. А в вышеописанной ситуации с молодой собакой можно ожидать, что животное бросится в погоню за "убегающим от нее" объектом.
У рыб зрительно-двигательные реакции имеют два проявления.
Одно заключается в движении глаз (оптокинетический нистагм), а другое - в движении всего тела (собственно оптомоторная реакция).
Зрительно-двигательная реакция имеет большое биологическое значение для рыб. Она позволяет максимально оптимизировать ориентировочное поведение животных. Эта реакция облегчает поиск пиши, контроль за источниками опасности, позволяет своевременно распознавать половых партнеров и пищевых конкурентов и, конечно, облегчает движение в составе многочисленной стаи.
Способность рыб держаться в определенном месте водоема с сильным течением также объясняется врожденной оптомоторной реакцией. В этом можно убедиться, если провести на рыбах опыт, схема которо-го показана на рис. 15.18.
При перемещении банки с рыбами навстречу потоку воды рыбы ориентируют тело по направлению перемещения окружающих предметов.
Если сосуд с рыбой оставить неподвижным в том же потоке воды, рыбы утрачивают ориентацию, так как предметы вокруг остаются неподвижными.
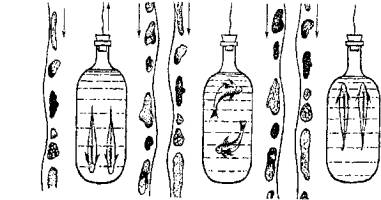
Рис. 15.18. Модель, демонстрирующая оптомоторную реакцию рыб в потоке воды
Наконец, направление движения сосуда совпадает с направлением потока воды, и рыба изменяет положение тела, ориентируясь на движение окружающих предметов.
Таким образом, ориентация рыбы в водном потоке определяется зрительно-двигательной реакцией.
Исключительно важное значение зрительно-двигательная реакция имеет для выклюнувшейся молоди. Шансы на выживание малька-одиночки крайне малы. Стайный образ жизни резко повышает шансы малька на выживание. Поэтому рефлекс следования за сородичами и импринтинг у высших позвоночных можно рассматривать как инструмент решения одной и той же биологической проблемы.
Небезынтересно заметить, что у экологически разных видов рыб зрительно-двигательные реакции проявляются неодинаково. Так, для донных рыб характерен оптокинетический нистагм. У пелагических рыб с хорошим зрением превалируют оптомоторные реакции.
Анализ поведения одиночных и стайных пелагических рыб показывает, что оптомоторные реакции - это атрибутика коллективных животных. Более того, степень проявления оптомоторных реакций у отдельных членов стаи зависит от численности стаи. Например, подсадка к одной верховке 5-7 рыбок значительно усиливает рефлекс следования. Дальнейшее увеличение численности еще более стабилизирует стаю, делает ее более монолитной, с быстрыми реакциями на стимул. Так реагируют на увеличение численности стаи типично стайные рыбы: уклея, гольян, вобла, сардина. Подобная картина характерна и для нестайных рыб, но в раннем возрасте.
Однако при формировании стаи из взрослых рыб, ведущих одиночный образ жизни или образующих в природе мелкие сообщества, стабилизации стаи не происходит. При объединении в большие стаи гуппи, ершей, щук оптомоторные реакции рыб ослабевают.
Таким образом, реакцию следования, возникающую у рыб на базе оптомоторных эффектов, можно рассматривать как предтечу импринтинга у высших позвоночных животных. Основным стимулом, запускающим оптомоторную реакцию у рыб, является оптическая сенсорика. В раннем онтогенезе оптомоторная реакция проявляется у всех без исключения видов рыб. Последующий переход к одиночному образу жизни ослабляет оптомоторную реакцию. В раннем онтогенезе она является инструментом использования филогенетического и онтогенетического коллективного опыта.
И. П. Павлов экспериментально разработал концепцию условных рефлексов. Используя его концепцию, можно объяснить многие аспекты научения и приобретения опыта. На базе врожденных рефлексов образуются условные рефлексы, обеспечивающие срочное приспособление животных к изменению среды. В свою очередь, условные рефлексы могут быть очень устойчивыми и в конце концов могут закрепляться наследственно, т. е. превращаться в безусловные. Скорость образования и устойчивость условного рефлекса существенным образом определяются экологической значимостью стимула и последствиями его сохранения в качестве врожденного рефлекса.
Важность экологической значимости стимула иллюстрируют опыты по выработке условного рефлекса на свет у экологически различающихся видов рыб. Например, условные оборонительные рефлексы у горбыля образуются после 6-8 сочетаний. У морского ерша - донной рыбы с плохим зрением - тот же условный рефлекс не образуется даже после 345 подкреплений. Условный рефлекс на звук у ската-хвостокола - обитателя мелководий - получали после 5-7 сочетаний, а у глубоководных скатов - после 25- 30 сочетаний условного и безусловного стимулов.
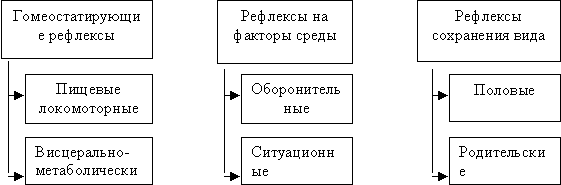
С позиции биологической значимости А. Д. Слоним разбил условные и безусловные рефлексы на несколько категорий (см. схему).
Современные исследователи поведения животных не ограничиваются рефлекторным подходом. Надо отметить, что сложные формы поведения животных не укладываются в рамки формулы "стимул - ответная реакция". Однако простые локомоторные акты (прежде всего спинальные) и регуляция вегетативных функций вполне вписываются в рамки рефлекторной теории. Вне всякого сомнения, в процессе научения и накопления опыта условные рефлексы играют важную роль.
Привыкание и сенсибилизация в процессе научения. После пересадки годовика карпа из зимовальных прудов в нагульные в ответ на появление на берегу трактора, развозящего комбикорм, рыба проявляет испуг и реакцию избегания. Повторное появление трактора на берегу вызывает менее выраженную реакцию избегания. Через несколько дней большая часть стаи перестает проявлять реакцию бегства в ответ на шум, производимый трактором. Рыба привыкает к данному раздражителю.
Привыкание - это понижение реактивности по отношению к стимулу. Скорость привыкания и его степень зависят от природы стимула, его силы, регулярности предъявления, а также от физиологического состояния животного. Так, использование того же нагульного пруда для водопоя стада коров вначале вызывает у рыб реакцию избегания. Регулярное появление стада на берегу пруда вызывает привыкание рыб. Причем ежедневное трехкратное появление коров на водопое сформирует привыкание скорее, чем ежедневное однократное или трехкратное нерегулярное (1-2 раза в неделю).
В то же время привыкание маловероятно в случае посещения водоема хищником, например выдрой. Привыкание не разовьется, даже если хищник будет появляться в водоеме регулярно и часто.
В последнем случае у карпа развивается явление, называемое сенсибилизацией. Сенсибилизация - это повышение реактивности в ответ на действие повторяющихся стимулов. Повторное предъявление рыбе стимула, сопряженного с опасностью для жизни, оценивается рыбой как нарастающая опасность по сравнению с единичным стимулом. Следовательно, в данной ситуации сенсибилизация имеет большую биологическую целесообразность по сравнению с привыканием.
Привыкание имеет вполне определенный биологический смысл, так как позволяет экономить энергию, которая потребуется для выполнения поведенческих актов в случае появления тривиальных раздражителей. Кроме того, привыкание усложняет хищникам поиск потенциальной жертвы во время ответа (карпа) на действие тривиального раздражителя. Привыкание можно рассматривать и как способ фильтрации менее значимых стимулов из внешней среды.
Нейрофизиологическая база привыкания состоит в утомлении рецепторного аппарата и (или) адаптации нейрона мозга к определенной афферентной информации. Возникновение привыкания связывают также и с ассоциативным обучением.
Знание физиологии привыкания имеет большое практическое значение в условиях искусственного разведения рыб. Любой технологический фактор может привести в стрессовое состояние рыбу с последующим ее отходом. Поэтому каждый новый технологический прием или внедряемый в производство механизм необходимо вводить осторожно, давая возможность рыбе выработать привыкание к нему.
Развитие поведения в онтогенезе. Оно имеет видовую специфичность. В то же время для всех видов рыб в раннем онтогенезе характерны одинаковые проблемы. Перед всеми мальками, поскольку они представляют легкую добычу для врагов, беззащитны перед изменениями окружающей среды, испытывают трудности с добыванием пищи в достаточном количестве и необходимого качества, стоит задача выжить. Проблемы молоди совсем не похожи на проблемы их родителей. Молодь имеет маленькие размеры тела, не способна к самообороне, привлекательна для хищников, которые не являются таковыми по отношению к их родителям, нуждается в особом (детском) питании.
Другая проблема для развивающейся молоди заключается в том, как измениться, чтобы превратиться в более или менее независимую взрослую особь.
Эволюционные изменения поведения молоди начинаются еще на личиночной стадии развития. В этот период молодь проявляет светотаксические и хемотаксические реакции, а также термические предпочтения.
Однако наиболее резкие количественно и качественно заметные изменения в поведении молоди происходят после перехода на активное питание. К этому моменту молодь большинства рыб собирается в стаи. При наличии пищи молодь растет быстро. У нее развиваются сенсорные системы, при помощи которых рыбки устанавливают связь с окружающим миром. Уже в недельном возрасте у мальков карпа наблюдаются исследовательское повеление, реакции избегания на зрительные, звуковые, химические, температурные и некоторые другие стимулы. На первом году жизни карпа его физические возможности невелики. Для выживания сеголетки держатся стаями, в которых численность особей составляет сначала тысячи, а к концу сезона сотни особей. Стаи держатся мелководий, которые хорошо прогреваются днем и в которых размножаются планктонные организмы. Молодь карповых является легкой добычей не только таких хищников, как Щука или окунь, но и других рыб - ротана, пескаря, голавля. Поэтому в естественных условиях выживаемость молоди карповых к зимовке не достигает 10 %. В рыбоводных специализированных прудах жизнь молоди более безопасна, поэтому выживаемость может превышать 95 %.
Зимует молодь в глубоких местах прудов, где вода не промерзает, формируя стаи из нескольких сотен особей. Во время зимовки молодняк не питается, да и в целом малоактивен.
С весенним теплом жизнь рыб активизируется. За период зимовки рыбы расходуют жировые запасы. Поэтому в весенне-летний период обостряется исследовательское поведение рыб, повышается их пищевая активность. Двухлетки начинают осваивать новые пищевые ниши. Если на первом году жизни карп проявляет себя как типичный планктонофаг, то на втором году жизни его пищевое поведение ориентировано в основном на бентос.
К концу лета при благоприятных условиях масса карпа возрастает до 500-700 г, что резко изменяет его зависимость от внешних факторов. Уменьшается количество его потенциальных врагов, но возникает проблема пищевых конкурентов. Поэтому стаи мельчают, а скорость перемещения рыб по водоему с целью поиска пищи возрастает. Фактически к концу второго года жизни поведение карпа стабилизируется на несколько лет. Оно обогащается только в связи с процессом репродукции. На 4-5-м году жизни карп достигает половой зрелости и у него проявляется половое поведение.
§60. ДВА ПОДХОДА К ИЗУЧЕНИЮ ПОВЕДЕНЧЕСКОГО АКТА
Проблема поведения животных привлекала внимание ученых на протяжении веков. Однако до сих пор степень понимания поведения животных (особенно сложных форм) не удовлетворяет специалистов. К настоящему времени известны два принципиально различных подхода к изучению проблемы поведения.
В основе первого направления лежит рефлекторная теория. Еще в начале XX века основоположники теории рефлексов И. П. Павлов, К. Шеррингтон и др. считали, что теория рефлексов удовлетворительно объясняет только спинальные ответы организма. Однако и в этой части не существует "чистых", обособленных рефлексов. Самая простая реакция организма на раздражитель охватывает не только тот узенький набор структур, который именуется "рефлекторной дугой" (рис. 15.19), но и целый комплекс центральных и периферических образований в пределах нервной системы и вне ее, которые, кроме того, взаимодействуют между собой.
Очевидно, что даже простые ответы на стимулы не живут отдельно сами по себе, а интегрируются с другими проявлениями нервной деятельности. И это относится как к низшим, так и к высшим позвоночным.
Рефлекторная дуга как морфологическая основа поведенческого акта изначально не выдерживала никакой критики. Любая ответная реакция организма на раздражение циклична. Голодная щука не откажется от поисков жертвы, если бросок на проплывающего перед ней карася оказался нерезультативным. Поиск или ожидание будут продолжаться до тех пор (даже если перед щукой нет раздражителя - карася), пока щука не удовлетворит свой аппетит.
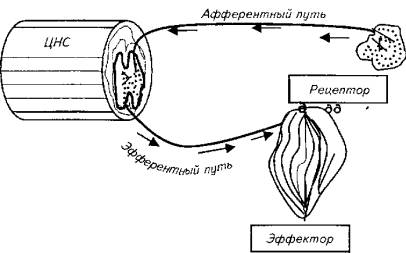
Рис. 15.19. Схема рефлекторной дуги
С другой стороны, раздражитель (карась) не вызовет ответного действия хищника, когда у того переполнен желудок. Раздражитель в паре "стимул - реакция" выполняет триггерную (пусковую) роль, когда есть потребность в данной реакции. Еще более наглядно об этом свидетельствуют половые отношения между потенциальными партнерами в нерестовый период и вне его.
Примат стимула в формировании различных поведенческих реакций и по сей день просматривается в методологических подходах при изучении поведения животных даже у таких выдающихся ученых в области этологии, как К. Лоренс, Дж. Кантор и др. Однако, невзирая на авторитеты, надо признать, что целостного понимания поведения животных эта методология не дает.
Другой принципиально отличный подход к изучению поведения животных постулирует, что моторная реакция животного - это внешнее проявление глубоких внутренних процессов, затрагивающих не только центральную нервную систему, но и метаболизм. Поведение животных диктуется биологической потребностью животного. Соматические реакции проявляются (или не проявляются) при возникновении внутренней мотивации поведения.
Такой подход в изучении поведения животных получил название теории функциональных систем. Основоположником данного направления исследований является П. К. Анохин. Его последователи во главе с К. В. Судаковым успешно развивают этот подход к проблеме поведения. Главная идея теории такова (рис. 15.20).
Внешний стимул только тогда спровоцирует поведенческий акт, когда в организме имеется потребность и раздражитель ассоциируется с возникшей биологической потребностью организма, т. е. если у щуки возникло чувство голода, то она совершит бросок на более мелкую рыбу, попавшую в ее поле зрения (востребованный раздражитель). Она не бросится на рыбу, превышающую ее собственные размеры, или на проплывающую мимо лодку (невостребованные раздражители). Специалисты называют это явление врожденным пусковым механизмом.
Результат броска влияет на потребность положительно или отрицательно (в случае неудачи). В первом случае щука прекращает охоту, во втором - будет повторять свои действия до удовлетворения пищевой потребности. При этом происходит постоянное сопоставление результата поведенческого акта с ожидаемым результатом, т. е. главным в данной ситуации является не раздражитель, а биологическая потребность - чувство голода.
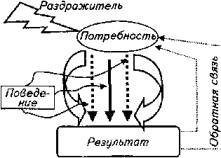
Рис. 15.20. Схема провокации поведенческого акта животного
внешним раздражителем
Таким образом, для функциональной системы пищевого поведения рыбы характерны цикличность блока стимул - ответ, наличие обратной связи между результатом поведения рыбы и потребностью; системообразующее значение биологической потребности и принцип саморегуляции системы. Раздражитель в этой схеме всего лишь триггер стереотипической реакции.
Важно понимать, как формируется биологическая потребность и возникает поведенческая мотивация у животного. Согласно теории функциональных систем потребность (чувство голода, жажды, чувство тревоги, опасности, дискомфорт при созревании икры и молок) развивается при изменении гомеостаза, который оценивается определенными показателями внутренней среды. Так, пищевая потребность возникает при изменении следующих гомеостатических констант:
снижении уровня глюкозы в крови; уменьшении пула жирных кислот; снижении осмотического давления крови; возникновении голодной моторики желудочно-кишечного тракта.
В ответ на изменение этих констант происходит возбуждение соответствующих рецепторных механизмов сосудистого русла, желудочно-кишечного тракта, других органов. В лимбической системе происходят анализ поступающей от рецепторного аппарата афферентации, афферентный синтез и созревает решение.
Многочисленные опыты на низших и высших позвоночных позволяют говорить о том, что у всех животных независимо от уровня их организации роль пейсмекера биологической мотивации принадлежит гипоталамусу.
Кора и подкорковые образования высших позвоночных имеют второстепенное значение и больше причастны к выработке тактики удовлетворения потребности (поведения). При разрушении корковых и подкорковых структур биологические мотивации поведения не исчезают. Разрушение ядер гипоталамуса при сохранении коры элиминирует мотивации.
Это же подтверждают и опыты по функциональному нарушению структур головного мозга. Так, аппликация атропина на передние отделы коры больших полушарий кошки лишь частично снижала чувство голода. При инъекции атропина в латеральную часть гипоталамуса чувство голода подавлялось полностью.
У рыб промежуточный мозг имеет еще большее значение в формировании поведенческих мотиваций, так как он в составе лимбической системы одновременно выполняет и интегрирующую функцию, которая у высших позвоночных ложится на кору больших полушарий и подкорковые структуры. В лимбическую систему рыб поступает афферентация от интеро-, проприо-, экстерорецепторов и от всех анализаторов - зрительного, слухового, обонятельного, осязательного, вкусового и от рецепторов боковой линии. Важную роль в формировании биологических мотиваций у рыб играют также средний, задний мозг и ретикулярная формация ствола.
В свою очередь, мотивационное возбуждение гипоталамуса повышает возбудимость и чувствительность рецепторного аппарата. Та же щука в голодном состоянии имеет обостренное зрение и функцию боковой линии, поэтому реагирует на малейшие колебания водной среды (потому и хватает блесну), идет на шум весла, а в темное время может подойти на свет фонаря.
Химическая основа мотивационного поведения изучена недостаточно. Однако известна причаст-ность к нему адренергических, холинергических веществ и дофамина. Эти вещества вызывают специфические изменения химических свойств отдельных нейронов мозга. В результате изменяется функциональная лабильность нервных клеток, повышается их способность к конвергенции афферентации и повышается общая чувствительность ядер гипоталамуса к действию "востребованных" раздражителей, удовлетворяющих биологическую потребность. Таким образом, возникает доминирующая мотивация, настраивающая ядра гипоталамуса на подкрепляющее воздействие.
У голодной щуки вне брачного сезона появление полового партнера только обостряет чувство голода. Во время брачного периода, наоборот, половая доминанта усиливается действием других раздражителей, например появлением потенциальной жертвы - карася.
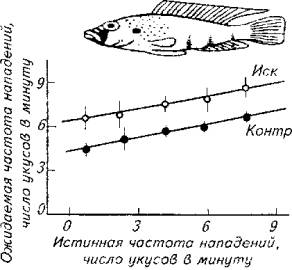
Рис. 15.21. Аддитивный эффект предъявления самца-соперника
(верхняя линия)
Изучение данного явления позволило сформулировать закон разнородной суммации, в соответствии с которым разнородные стимулы создают аддитивный эффект в формировании поведенческой реакции. Наглядно этот закон иллюстрирует опыт на агрессивных самцах цихлидовых рыб (рис. §5.21).
Взрослого самца помещали в один аквариум с молодью. Самец периодически совершает броски на молодых рыбок (нижняя линия на рис. 15.21). После того как за стеклом помещали куклу самца того же вида, агрессивность самца по отношению к молоди возросла (верхняя линия на рис. 15.21). Аддитивный эффект наблюдается сразу после предъявления соперника, а затем возвращается к исходному уровню. Степень аддитивного влияния зависит еще и от окраски самца-соперника.
В этих условиях проявляются основные свойства доминанты - способность к суммации, стойкость, инертность.
Возникновение доминанты возбуждения гипоталамуса сопровождается постановкой цели и формированием акцептора результата действия в структурах лимбической системы. Доминанта выбирает и соответствующий тип поведения, т. е. механизм эффекторной реализации поведения.
Таким образом, структурным элементом поведения выступает функциональная система - набор центральных и периферических нервных образований и эффекторных механизмов, действующих согласованно до получения полезного эффекта (удовлетворения потребности), т.е. замкнутая, циклическая и саморегулируемая система.
Мотивационное состояние животного возникает как результат взаимодействия двух факторов: 1) внутренней и 2) внешней среды. В свою очередь, наблюдаемое поведение есть некий оптимизированный процесс взаимодействия физиологического состояния животного и афферентного потока, возникшего в конкретной пространственно-временной ситуации. Данный фундаментальный тезис поясняет следующий пример. Щука может проявить одинаковую пищевую (поисковую) активность при:
I) сильном голоде (внутреннее состояние) и сомнительной доступности жертвы (слабый внешний стимул);
2) слабом голоде и наличии доступной пиши (сильный внешний стимул).
Зависимость между мотивационным состоянием животного и поведением не прямая. На рис. 15.22 предпринята попытка графической и математической оценки этих двух явлений на примере проявления полового поведения самца гуппи.
Анализ диаграммы показывает, что максимальную активность слабоокрашенный самец гуппи проявляет в ситуации, когда видит крупную (созревшую) самку. С другой стороны, в отношении одной и той же незрелой самки наиболее активен самец, у которого наиболее яркая окраска (т. е. он находится в состоянии наивысшего полового возбуждения). Между этими двумя крайностями есть промежуточные варианты. Максимум активности приходится на вариант, в котором внутренние и внешние стимулы имеют максимальное напряжение (верхняя кривая).
Кроме того, поведение животного может быть результатом предвидения последствий одного или другого рода активности. Поведение формируется как компромисс между потребностью и затратами энергии при достижении результата. Очень голодная щука может и не броситься на жертву, если та мала и неуловима. Однако даже сытая щука может совершить бросок, если добыча привлекает ее размером и доступностью.
Совершенно очевидно, что поведенческая реакция есть результат внутренней мотивационной компоненты и афферентационного синтеза.
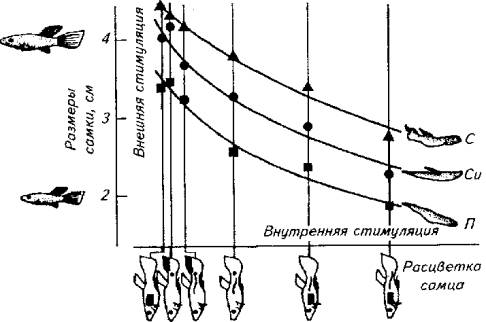
Рис. 15.22. Взаимосвязь внутреннего состояния (яркость окраски тела самца) и силы внешней стимуляции (размер самки) при формировании поведения ухаживания самца за самкой

Рис. 15.23. Смешенная активность (колюшка вентилирует гнездо без икры)
Справедливости ради надо признать, что есть образцы даже простого поведения животных, плохо согласующиеся с постулатом биологической потребности. Примером является так называемая смещенная активность. Под этим термином скрывается поведение животного, не соответствующее обстановке и биологической потребности самого животного. Например, перед началом нереста самец трехиглой колюшки совершает ритуальные движения ухаживания. Однако он может внезапно оставить самку и переключиться на вентиляцию гнезда, что до появления в нем икры лишено смысла (рис. 15.23).
Однако о причинах смещенной активности ведутся споры. Тинберген видел причину в конфликте двух биологических мотиваций, имеющих исключительно большое значение. Избыточная энергия первой мотивации (спаривание) разряжается за счет второй (ухаживание за гнездом).
Подобным образом разрешаются и конфликтные ситуации. Вблизи построенного гнезда (своя территория) самец колюшки активно нападает на чужака, вторгшегося на его территорию. Попав на чужую территорию, тот же самец не нападает на хозяина, а активно ретируется на свою территорию.
Однако, если самцы встречаются на границе владений, конфликт разрешается иначе. Они проявляют смещенную активность. На границе участков самцы принимают позы копательного поведения, что с точки зрения целесообразности лишено всякого смысла (рис. 15.24).
В данной ситуации две сильные мотивации (атаковать или бежать?) разряжаются во внешне бессмысленном копательном поведении.
Не совсем понятен и механизм выбора приоритетности мотиваций. Одновременно в организме рыбы формируется несколько потребностей и, следовательно, несколько мотиваций. В течение одного дня животные осуществляют различные виды полезного поведения. В более или менее константных условиях жизни у животного складывается определенный распорядок, в соответствии с которым каждый мотивированный потребностью вид деятельности занимает определенное время суток. Остается неясным, каким образом происходит распределение времени по видам активности. Заслуживает специального изучения и механизм перераспределения времени по видам активности в случае появления дополнительного сильного биологически значимого фактора.

Рис. 15.24. Копательная поза (смешенное поведение) агрессивных самцов, встретившихся на границе своих территорий
Д. Мак-Фарленд (1988) ввел в этологию животных понятие упругости поведения, которое несколько проясняет ситуацию.
Упругость поведения - это мера того, до какой степени может быть сжат во времени любой вид активности животного под влиянием других видов. Данное явление характерно для всех животных независимо от уровня их эволюционного положения.
Рыбы не являются исключением из этого правила. Время, затрачиваемое, например, парой лялиусов на исследовательскую деятельность, пищевую активность и игровое поведение, перераспределяется по мере созревания самки. Чем ближе момент нереста, тем меньше времени уделяется игровому поведению. В определенный момент в поведении самца появляются новые виды активности: ухаживание за самкой и построение гнезда. Параллельно ограничивается время на другие виды активности. После нереста самец сокращает время на питание, исследовательскую активность, игры до нуля. Весь суточный запас времени в этот период занимает активность, связанная с ухаживанием и охраной гнезда. В других ситуациях более эластичным оказывается пищевое поведение.
Эластичность поведения иллюстрирует и следующий опыт на бойцовых рыбках (рис. 15.25).
У бойцовой рыбки вырабатывали условный рефлекс на прохождение через специальный тоннель, помещенный в аквариум.
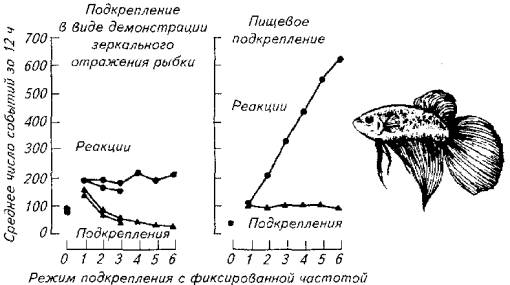
Рис. 15.25. Эластичность агрессивного поведения (слева) и эластичность пищевого поведения у бойцовой рыбки
Появление рыбки в тоннеле подкрепляли зеркальным отражением той же особи, чем провоцировали агрессивность самца. В другом варианте опыта использовали пищевое подкрепление.
Как следует из диаграммы ответов, агрессивная активность оказалась неэластичной в отличие от пищевой активности рыб. В другом варианте время, отводимое на пищевое поведение, расширялось по мере увеличения частоты подкрепления ответных реакций. Количество проходов через тоннель в течение 12ч наблюдений при подкреплении зеркальным отражением не зависело от частоты предъявляемого подкрепления.
По принципу функциональной системы осуществляется не только регуляция поведения рыб (набор моторных реакций), но и поддержание гомеостаза. Константы гомеостаза делят на жесткие и мягкие (изменяющиеся в более широких пределах). Для рыб жесткими константами гомеостаза являются уровень кислорода в крови, осмотическое давление внутренней среды. В широких же пределах изменяются температура тела, давление крови, уровень глюкозы в крови, концентрация сывороточных белков и жирных кислот в крови.
В качестве примера рассмотрим функциональную систему постоянства кислорода в крови. Это гомеостатическая функциональная система с активным внешним звеном саморегуляции.
Результатом действия данной системы является поддержание кислорода на определенном уровне. Его изменения возбуждает (угнетает) дыхательный центр (группа нейронов спинного, продолговатого, промежуточного мозга). Возникающее возбуждение поступает в гипоталамус, где в результате афферентного синтеза формируется программа нормализации уровня кислорода в крови, которая включает в себя соматический (поведенческий) и вегетативный механизмы. Они запускаются напрямую из гипоталамуса или через влияния по нисходящим нервным путям (рис. 15.26).
Соматический ответ заключается в том, что рыба ищет воду с высоким содержанием кислорода или поднимается к поверхности и жадно хватает атмосферный воздух.
Вегетативная регуляция сводится к целому спектру влияний на жаберный аппарат (повышаются частота дыхания, проницаемость через жаберные мембраны кислорода), сердечно-сосудистую систему (возрастает частота сердечных сокращений, выбрасываются в русло депонированные эритроциты). Происходят адаптационные изменения в метаболизме рыбы, включается гормональная регуляция вспомогательных вегетативных процессов. Дополнительно кислород перекачивается из плавательного пузыря, включаются в газообмен кожа и желудочно-кишечный тракт.
И соматические, и вегетативные влияния приводят к одному результату: количество поступающего в организм рыбы кислорода возрастает, его потребление снижается. Акцепторы результата информируют гипоталамус о произошедших изменениях. Если полезный приспособительный эффект достигнут, регулируемая функция гипоталамуса тормозится. Если результат не удовлетворяет лимбическую систему и поставленная цель не достигнута, процесс соматовегетативной регуляции повторяется.
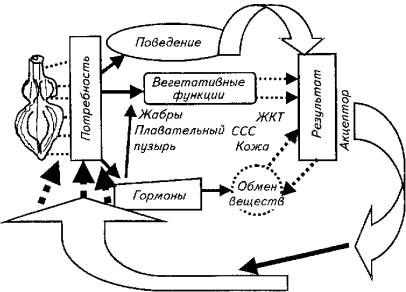
Рис. 15.26. Функциональная система поддержания количества кислорода в крови рыб
Обратная связь осуществляется посредством гуморального канала (О2, СО2, рН-рецепторы гипоталамуса оценивают состав крови непосредственно) и нервной афферентации от соответствующих рецепторов периферических органов.
Таким образом, данная функциональная система хотя и относится к гомеостатическому типу, но включает все возможные эффекторные рычаги регуляции - поведенческий, вегетативный, метаболический.
В заключение хочется еще раз подчеркнуть, что теория функциональных систем дает возможность исследователю объяснить цельное поведение рыбы независимо от того, сколь сложна биологическая задача (потребность).
Эта теория применяется для объяснения не только поведения отдельных индивидуумов, но и для трактовки стайного (группового) поведения рыб.
Формирование групповых отношений определяется врожденными причинами, которые закрепляют биологическую целесообразность группового образа жизни. Так, трудно представить стайный образ жизни крупных хищников. С другой стороны, мало шансов на выживание в одиночестве у верховки, уклеи. Изоляция от группы уклеи приводит ее в дискомфортное, стрессовое состояние, из которого рыба старается выйти как можно быстрее, объединившись со стаей.
В стае существуют сложные иерархические отношения; каждая особь в стае занимает определенное место и свою специфическую групповую функцию. Правда, эта позиция индивидуума не прочна; она постоянно подвергается переоценке. Каждый член стаи старается поднять свой статус и занять более выгодное положение.
Структура стаи у разных видов рыб может быть различной, Однако в стае с лидером есть позиции более и менее выгодные, что и побуждает особей к постоянным иерархическим отношениям. Так, менее выгодно положение в конце стаи: последним меньше достается корма, а в случае преследования они первые кандидаты в жертвы хищника. Объединение в стаю выгодно ее членам: в стае им комфортно и безопасно (менее опасно).
В свою очередь, стая полезна для популяции. Стая удовлетворяет СБОИ пищевые, оборонительные, репродуктивные функции и потребности популяции вида в целом. Стая значительно эффективнее адаптируется к изменениям среды обитания, так как, с одной стороны, индивидуальный положительный опыт становится достоянием всех членов стаи, а с другой - стая обеспечивает проверенным временем полезным групповым опытом каждого члена стаи.
В природе постоянно происходит некоторое перемешивание групп, особенно заметное при миграциях молоди. Последнее также полезно для популяции, так как сводит к минимуму вероятность инбридинга. Кроме того, стая стабильна тогда, когда в ней достаточное количество особей.
Однако полезным результатом функциональной системы группового поведения все равно является равновесное состояние стаи. Стая тем стабильнее, чем большее количество ее членов стереотипично и синхронно реагируют на дестабилизирующий стаю фактор.
Функциональная система поведения стаи складывается из нескольких биологически детерминированных поведенческих блоков:
системы пищевого поведения;
системы оборонительного поведения;
системы игрового поведения;
системы полового поведения;
системы ориентировочного поведения,
Структурированная стая наиболее эффективно достигает полезно-приспособительного результата.
Таким образом, как индивидуальное, так и стайное поведение рыб нацелено на удовлетворение определенных биологических потребностей. Использование стайного или индивидуального образа жизни рыбами - это всего лишь способ решения определенной биологической задачи.
From: www.fishpatholog.amillo.net/Fiziologia/Fiziologia.htm
Дата добавления: 2014-12-11 | Просмотры: 1851 | Нарушение авторских прав
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 34 | 35 |
|