|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология
|
Характеристики красной крови сеголетков и годовиков карпа
| Hb, %
| Гематокрит, %
| Количество эритроцитов, млн
| НЬ в одном эритроците, мг%
| Объем эритроцита, мкм
| | 10,8*
| 35,8*
| 1,24*
| 86/32*
| 292*
| | 9,7**
| 32,8**
| 1,20**
| 86/32**
| 274**
| * Сеголетки (октябрь). ** Годовики (май).
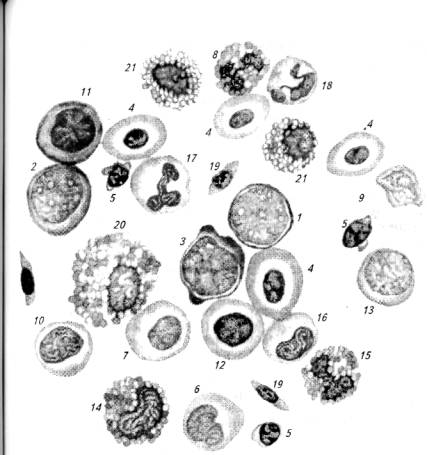
Рис. 6.1. Клетки крови осетра:
1-гемоцитобласт; 2- миелобласт; 3- эритробласт; 4- эритроциты; 5- лимфоциты; 6- моноцит; 7- нейтрофильный миелоцит; 8- сегментоядерный эозинофил; 9- монобласт; 10- промиелоцит; 11 - базофильный нормобласт; 12- полихроматофильный нормобласт; 13- лимфобласт; 14- эозинофильный метамиелоцит; 15- палочкоядерный эозинофил; 16- профильный метамиелопит; 17- палочкоядерный кейтрофил; 18- сегментоядерный нейтрофил; 19 - тромбоциты; 20- эозинофильный миелоцит; 21 - клетки с вакуолизированной цитоплазмой
Характеристика красной крови зависит от факторов внешней среды. Обеспеченность рыбы гемоглобином определяется температурой воды. Выращивание рыбы в условиях пониженного содержания кислорода сопровождается увеличением общего объема крови, плазмы, что повышает эффективность газообмена.
Характерной особенностью рыб является полиморфизм красных
- одновременное присутствие в кровяном русле эритроцитных клеток различной степени зрелости (табл. 6.8).
6.8. Эритроцитарный ряд форели (%)
| Длина рыбы, см
| Незрелые формы эритроцитов
| Зрелые эритроциты
| | эритробласт
| нормобласт
| базофильный
| полихромофильный
| | 4,2
| 1,5
| 4,1
| 11,8
| 19,5
| 63,1
| | 7,6
|
| 4,4
| 5,7
| 15,3
| 74,6
| | 18,2
|
| 2,2
| 7,7
| 12,3
| 77,8
|
Увеличение количества незрелых форм эритроцитов связано с сезонным усилением обмена веществ, кровопотерями, а также с возрастными и половыми особенностями рыб. Так, у производителей наблюдается 2-3-кратное увеличение незрелых эритроцитов по мере созревания гонад, достигающее 15 % у самцов перед нерестом.
В эволюции красных клеток крови рыб выделяют три этапа, каждый из которых характеризуется образованием морфологически довольно самостоятельных клеток - эритробласта, нормобластов и собственно эритроцита.
Эритробласт является самой незрелой клеткой эритроидного ряда. Эритробласты рыб можно отнести к средним и крупным клеткам крови, так как их размеры составляют от 9 до 14 мкм. Ядро этих клеток имеет красно-фиолетовый цвет (в мазке). Хроматин равномерно распределяется по ядру, образуя сетчатую структуру. При большом увеличении в ядре можно обнаружить от 2 до 4 ядрышек. Цитоплазма этих клеток резко базофильна. Она образует сравнительно правильное кольцо вокруг ядра.
Базофильный нормобласт образуется из эритробласта. Эта клетка имеет более плотное ядро меньшего размера, которое занимает Центральную часть клетки. Цитоплазма характеризуется слабовыраженными базофильными свойствами.
Полихроматофилъный нормобласт отличается еще меньшим, с резко очерченными краями ядром, которое несколько смещается от центра клетки. Другой его особенностью является то, что ядерный хроматин располагается радиально, образуя довольно правильные сектора в пределах ядра. Цитоплазма клеток в мазке имеет не базофильное, а грязно-розовое (светло-сиреневое) окрашивание.
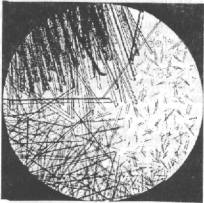
Рис. 6.2. Кристаллы гемоглобина рыб
Оксифильный нормобласт имеет округлую форму с центрально расположенным округлым и плотным ядром. Цитоплазма располагается широким кольцом вокруг ядра и имеет хорошо различимую розовую окраску.
Эритроциты рыб завершают эритроидный ряд. Они имеют овальную форму с повторяющим их форму плотным ядром красно-фиолетового цвета. Хроматин образует скопления в виде специфических глыбок. В целом зрелый эритроцит похож на оксифильный нормобласт как по характеру окраски ядра- и цитоплазмы в мазке, так и по микроструктуре протоплазмы. Его отличает лишь вытянутая форма.
Скорость оседания эритроцитов (СОЭ) у рыб в норме составляет 2-10 мм/ч.
Белые клетки крови (лейкоциты). Лейкоциты крови рыб представлены в большем количестве, чем таковые у млекопитающих. Для рыб характерен лимфоцитарный профиль, т. е. более 90 % белых клеток составляют лимфоциты (табл. 6.9, 6.10).
6.9. Количество лейкоцитов в 1 мм3
| Вид рыб
| Количество клеток, тыс/мм2
| Вид рыб
| Количество клеток, тыс/мм2
| | Форель
|
| Щука
| 30-110
| | Карп:
|
| Толстолобик
| 60-100
| | сеголетки
| 50-80
| Осетр
| 15-30
| | годовики
| 80-160
|
|
| | двухлетки
| 30-120
|
|
| | трехлетки
| 60-90
|
|
|
6.10. Лейкоцитарная формула, %
| Вид и масса рыб, г
| Лимфоциты
| Моноциты
| ПМЯ клетки
| Эозинофилы
| Нейтрофилы
| | Карп:
0,26
|
|
| 0,1
| -
| -
| |
|
|
|
| 0,1
| 0,1
| |
|
|
|
| -
| -
| |
|
|
| 0,1
| -
| -
| | Форель:
|
|
|
|
|
| |
|
|
| 0,3
| -
| -
| |
|
|
| 0,5
| -
| -
| | Толстолобик
|
|
|
|
| 4,5
|
Фагоцитирующими формами являются моноциты и полиморфноядерные клетки. На протяжении жизненного цикла лейкоцитарная формула меняется под влиянием факторов внешней среды. Во время нереста снижается количество лимфоцитов в пользу моноцитов и полиморфноядерных клеток.
В крови рыб присутствуют полиморфноядерные клетки (гранулоциты), находящиеся на разных стадиях зрелости. Родоначальником всех гранулоцитов следует рассматривать миелобласт (рис. 6.3).
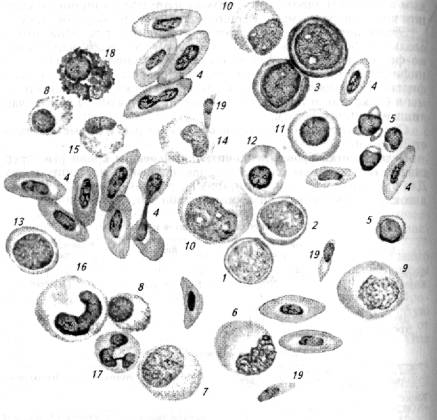
Рис. 6.3. Клетки крови карася:
1 - гемоцитобласт; 2- миелобласт; 3 - эритробласт; 4-эритроциты; 5 -лимфоциты; 6- моноцит; 7- нейтрофильный миелоцит; 8- псевдоэозинофильный миелоцит; 9- монобласт; 10- промиелоцит; 11 - базофильный нормобласт; 12 -- полихроматофильный нормобласт; 13 - лимфобласт; 14- нейтрофильный метамиелоиит; 15- псевдоэозинофильный метамиелоцит; 16- палочколдерный нейтрофил; 17 - сегментоядерный нейтрофил; 18- псевдобазофил; 19- тромбоцит
Эта клетка отличается крупными размерами и большим ядром красно-фиолетового цвета, которое занимает большую ее часть. Размеры миелобластов колеблются от 12 до 20 мкм. Микроструктура клеток характеризуется обилием рибосом, митохондрий, а также интенсивным развитием комплекса Гольджи. При созревании миелобласт переходит в промиелоцит.
Промиелоцит сохраняет размеры своей предшественницы, т.е. является крупной клеткой. По сравнению с миелобластом промиелоцит имеет более плотное ядро красно-фиолетового цвета с 2-4 ядрышками и слабобазофильную цитоплазму зернистой структуры. Кроме того, в этой клетке меньше рибосом.
Миелоцит мельче предыдущих клеток (10-15 мкм). Плотное круглое ядро утрачивает ядрышки. Цитоплазма занимает больший объем, имеет ярко выраженную зернистость, которая выявляется кислыми, нейтральными и основными красителями.
Метамиелоцит отличается ядром вытянутой формы с пятнистым хроматином. Цитоплазма клеток имеет неоднородную гранулярную структуру.
Палочкоядерный гранулоцит представляет собой дальнейший этап эволюции гранулоиитов. Отличительным признаком его является форма плотного ядра. Оно у него вытянутое, с обязательным перехватом. К тому же ядро занимает меньшую часть объема клетки.
Сегментоядерный гранулоцит представляет конечную стадию созревания миелобласта, т.е. является наиболее зрелой клеткой гранулярного ряда крови рыб. Его отличительной особенностью является сегментированное ядро. В зависимости от того, какой краской окрашиваются гранулы цитоплазмы, сегментоядерные клетки дополнительно классифицируют на нейтрофилы, эозинофилы, базофилы, а также на псевдоэозинофилы и псевдобазофилы. Некоторые исследователи отрицают наличие базофильных форм гранулоцитов у осетровых рыб.
Полиморфизм клеток отмечается и у лимфоцитов крови рыб. Наименее зрелой клеткой лимфоидного ряда считают лимфобласт, формирующийся из гемоцитобласта.
Лимфобласт отличается крупным округлым ядром красно-фиолетового цвета с сетчатой структурой хроматина. На долю цитоплазмы приходится узкая полоска, окрашиваемая основными красителями. При изучении клетки под большим увеличением обнаруживается много рибосом и митохондрий на фоне слабого развития комплекса Гольджи и эндоплазматического ретикулума.
Пролимфоцит представляет собой промежуточную стадию развития клеток лимфоидного ряда. От предшественника пролимфоцит отличается структурой хроматина в ядре: он утрачивает сетчатое строение.
Лимфоцит имеет красно-фиолетовое ядро различной формы (округлое, овальное, палочковидное, дольчатое), которое располагается в клетке асимметрично. Хроматин распределен в пределах ядра неравномерно. Поэтому на окрашенных препаратах в пределах ядра видны облаковидные структуры. Цитоплазма располагается асимметрично относительно ядра и часто образует псевдоподии, что придает клетке амебовидную форму.
Лимфоцит рыб - мелкая клетка (5-10 мкм). При микроскопировании мазков крови лимфоциты можно спутать с другими мелкими клетками крови- тромбоцитами. При их распознавании следует учитывать различия в форме клеток, ядра и границ распределения цитоплазмы вокруг ядра. К тому же и окрашенность цитоплазмы у этих клеток неодинакова: у лимфоцитов она синяя, у тромбоцитов -- розовая.
В свою очередь, лимфоциты крови - неоднородная группа клеток, различающихся по морфофункциональным признакам. Здесь довольно упомянуть о том, что выделяют Т- и В-лимфоциты, которые имеют неодинаковое происхождение и свои собственные уникальные функции в реакциях клеточного и гуморального иммунитета.
Моноцитоидный ряд белой крови рыб представляют, по крайней мере, три типа довольно крупных (11 - 17 мкм) клеток.
Монобласт является наименее зрелой клеткой этого ряда. Он выделяется крупным ядром красно-фиолетового цвета неправильной формы: бобовидной, подковообразной, серповидной. Клетки имеют широкий слой цитоплазмы со слабобазофильными свойствами.
Промоноцит отличается от монобласта более рыхлой структурой ядра и хроматином дымчатого вида (после окрашивания). Неравномерно окрашивается и цитоплазма этих клеток, отчего приобретает дымчатость.
Моноцит - наиболее зрелая клетка ряда. Имеет крупное ядро красно-фиолетового цвета с относительно небольшим количеством хроматинового вещества. Форма ядра чаще неправильная. На окрашенных препаратах цитоплазма сохраняет дымчатость.
Ухудшение условий содержания рыбы (гипоксия, бактериальная и химическая загрязненность водоема, голодание) приводит к увеличению фагоцитирующих форм. В процессе зимовки карпа отмечают 2-16-кратный рост количества моноцитов и полиморфноядерных клеток при одновременном уменьшении на 10-30 % количества лимфоцитов. Таким образом, за физиологическую норму следует принимать показатели рыб, выращенных в хороших условиях.
Тромбоциты крови рыб. Нет более противоречивой информации о морфологии и происхождении клеток крови, чем сведения о тромбоцитах рыб. Отдельными авторами существование этих клеток вообще отрицается. Однако более убедительной выглядит точка зрения о большом морфологическом разнообразии и высокой изменчивости тромбоцитов в организме рыб. Не последнее место в этом споре занимают особенности методических приемов при исследовании тромбоцитов.
В мазках крови, сделанных без применения антикоагулянтов, многие исследователи обнаруживают, как минимум, четыре морфологические формы тромбоцитов - шиловидную, веретенообразную, овальную и округлую. Овальные тромбоциты внешне практически неотличимы от мелких лимфоцитов (рис. 6.4).
Поэтому при подсчете тромбоцитов в мазке крови их количественная характеристика в 4 %, вероятно, занижается при использовании данной методики.

Рис. 6.4. Тромбоциты крови рыб
Известна и другая крайность. Некоторые авторы 82-95 % окрашенных по Романовскому клеток причисляют к тромбоцитам.
Более совершенные методы, например иммунофлюоресцентный со стабилизацией крови гепарином, позволили определить соотношение лимфоциты: тромбоциты как 1: 3. Концентрация тромбоцитов в 1 мм3 при этом составила 360 000 клеток.
Остается открытым вопрос о происхождении тромбоцитов у рыб. Распространенная точка зрения о едином с лимфоцитами происхождении из мелких лимфоидных гемобластов в последнее время подвергается сомнению. Ткань, производящая тромбоциты, у рыб не описана. Однако обращает на себя внимание то, что в отпечатках от срезов селезенки практически всегда обнаруживается большое количество овальных клеток, сильно напоминающих овальные формы тромбоцитов. Следовательно, есть основания полагать, что тромбоциты рыб образуются в селезенке.
Таким образом, можно определенно говорить о существовании тромбоцитов в классе рыб de facto, отметив при этом их большое морфологическое и функциональное разнообразие.
Количественная характеристика этой группы клеток не отличается от таковой у других классов животных.
Среди исследователей крови рыб существует единая точка зрения относительно функциональной значимости тромбоцитов. Подобно тромбоцитам других классов животных у рыб они осуществляют процесс свертывания крови.
У рыб время свертывания крови - довольно нестабильный показатель, который зависит не только от способа взятия крови, но и от факторов внешней среды, физиологического состояния рыбы (табл. 6.11).
Дата добавления: 2014-12-11 | Просмотры: 1447 | Нарушение авторских прав
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 34 | 35 |
|