|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология
|
Глава 3. НЕРВНАЯ СИСТЕМА РЫБ
Нервная система животного возникла в процессе его эволюции как инструмент, специализированный на объединении огромного количества структур многоклеточного организма в единое целое.
Важнейшая задача нервной системы заключается в поддержании постоянной связи животного со средой обитания и поддержании гомеостаза организма, адекватного изменениям, происходящим во внешней среде.
С позиции этологии можно рассматривать нервную систему как аппарат хранения опыта (исторического и личного) и инструмент научения под влиянием стимулов из внешнего мира.
Реактивность организма к факторам среды существовала и в донервный период. Так, у простейших, например амебы или инфузории туфельки, существует реактивность по отношению к изменениям среды (температуры, химического состава, освещенности). Однако их реакция на стимул чаще всего избыточно велика. Из-за отсутствия специализированных органов рецепции простейшие вынуждены реагировать на стимул всем организмом, т. е. с позиции многоклеточного животного неадекватно, с чрезмерными затратами энергии и (что не менее важно) времени.
Появление нервной системы в процессе эволюционного развития позволило животным оптимизировать свои отношения со средой обитания, сделать реакции организма более экономными и биологически более эффективными. Появление "многоклеточности" неизбежно привело к формированию нервной системы. Для слаженной работы многоклеточного организма химической регуляции недостаточно: она медлительна, требует больших затрат энергии и жестких стабильных условий (рН. температура, содержание кислорода).
У многоклеточных животных регуляцию физиологических функций осуществляет сложная система нейрогуморальных механизмов. Гуморальная (химическая) регуляция досталась им от низших форм жизни. Нервная регуляция - собственное эволюционное приобретение многоклеточных, обусловившее быстрое эволюционно-адаптивное развитие многоклеточных животных. Эволюция сохранила химическое звено, поскольку в определенных ситуациях химическая регуляция незаменима. Это прежде всего касается метаболической регуляции, регуляции процессов хранения генетической информации, дифференцировки клеток и тканей, органогенеза, иммунной защиты животного.
Основные свойства нервных клеток - чувствительность, раздражимость и возбудимость -позволяют нервной системе чутко реагировать на изменения среды, анализировать стимулы, оценивать со-стояние собственного организма и быстро принимать оптимальные решения при крайне малых затратах энергии. Нервная система призвана обеспечивать срочную регуляцию.
И. П. Павлов создал учение о нервизме, в соответствии с которым каждая клетка организма независимо от того, в составе соматического или висцерального органа она находится, является подконтрольной нервной системе. В то же время он подчеркивал, что между гуморальной и нервной регуляцией сохраняется тесная многоуровневая связь даже у высших позвоночных животных. Однако чаще всего в этом гармоничном ансамбле регуляторных механизмов лидирующую позицию занимает нервная система.
В своем эволюционном развитии нервная система прошла путь от малоэффективной диффузной структуры до сложнейшего трубчатого образования, строение и функции которого еще долго будут оставаться предметом напряженного изучения человеком.
Это подтверждают наблюдения за гидростатическими реакциями рыб, а также специальные опыты с применением электрофизиологических методов и методик с выработкой условных рефлексов на изменение гидростатического давления в искусственных водоемах.
Сопоставление размеров головного мозга и всего тела животных показало, что у рыб головной мозг менее развит, чем у наземных позвоночных. В ряду рыбы - амфибии -рептилии различия невелики. Однако при сравнении головного мозга рыб с головным мозгом птиц и особенно млекопитающих выявляется огромная разница.
Головной мозг рыб отличается не только размерами, но и уровнем организации. Он состоит из заднего, среднего и переднего отделов. Степень их развития различна и порой зависит не от эволюционного положения рыбы, а от ее экологической ниши
(рис. 3.1). Например, мозжечок более развит у акулы, чем у карася.
Анализ диаграммы 3.1 доказывает зависимость развития той или иной части головного мозга у костистых рыб от занимаемой ими экологической ниши и этологической характеристики. Так, передний мозг наиболее развит у угря, продолговатый - у сазана и язя. Средний мозг занимает больший удельный вес у активных рыб с острым зрением. У сома более развит мозжечок.
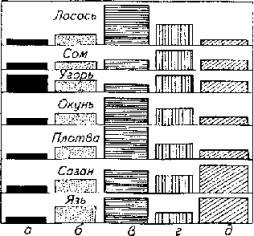
Рис. 3.1. Этологический анализ строения головного мозга костистых рыб:
а - обонятельные луковицы; 6 - передний мозг; в - средний мозг; г - мозжечок; д - продолговатый мозг (по М.Никитенко)
Фактическое отсутствие больших полушарий и слабое морфологическое обособление промежуточного мозга у рыб означает перераспределение функций между отделами головного и спинного мозга. Так, высшую интеграционную функцию у рыб выполняют средний, промежуточный мозг, мозжечок и продолговатый мозг. Интегратором сложных локомоторных реакций у рыб является спинной мозг.
Необходимо подчеркнуть, что спинной мозг рыб вообще проявляет большую степень автономности и имеет причастность к регуляции практически всех без исключения функций организма (локомоторных, висцеральных, метаболических). Единственное, что ускользает от его контроля, это сенсорная афферентация, афферентный синтез и механизм принятия решений на его основе.
Глубокому и всестороннему пониманию той или иной физиологической функции помогает анализ филогенетического развития обеспечивающей эту функцию системы органов. С этой позиции большой интерес представляет изучение морфофункциональных особенностей нервной системы у первичных хордовых, например у ланцетника.
§11. НЕРВНАЯ СИСТЕМА ЛАНЦЕТНИКА
Нервная система ланцетника - яркий пример ароморфоза, обеспечившего качественный эволюционный скачок в развитии животных организмов. Сейчас бесчерепные хордовые представлены единичными видами. Однако в свое время они дали начало такому идеоадаптационному изменению нервной системы, как цефализация, позволившему сохраниться и дойти до наших дней целому классу водных животных.
Что представляет собой нервная система ланцетника? Эмбриогенез ланцетника свидетельствует о том, что центральная нервная система позвоночных животных образуется за счет качественно нового материального источника - эктодермы. У беспозвоночных предшественник нервной системы другой - субэпителиальные чувствительные клетки.
Важной особенностью нервной системы ланцетника является неоднородность. Нервный тяж ланцетника представлен передним участком, условно называемым "головным мозгом", и каудальным участком, условно называемым "спинным мозгом". Поэтому деление центральной нервной системы бесчерепных многими исследователями не принимается, поскольку дифференциация краниальной и каудальной частей мозга выражена неярко. Действительно, нервный тяж ланцетника представляет собой диффузное образование, не имеет деления на дорзальные и вентральные рога.
Однако при рассечении тела ланцетника пополам только передняя половина сохраняет реактивность к механическим раздражителям. Задняя половина утрачивает чувствительность и подвижность. Таким обра-зом, головной конец мозга оказывает регулирующее влияние, по крайней мере, на двигательные рефлексы ланцетника.
Собственно нервных центров в мозге ланцетника не обнаружено. Их функции выполняют гигантские нервные клетки, которые располагаются в областях между 6-м и 11-м сегментами, а также между 39-м и 61-м сегментами. Указанные сегменты имеют по одной гигантской клетке. От нее отходит толстое волокно, которое тянется через другие сегменты. Считают, что эти биполярные клетки осуществляют одновременно связующую, координационную и проводниковую функции.
Интересно, что мозг ланцетника проявляет высокую чувствительность к свету. Гистологические исследования мозга выявили большое количество глазков, состоящих из пары клеток: крупной ганглиозной и покрывающей ее пигментной клетки. Светочувствительные глазки располагаются вдоль всего нервного тяжа, причем их концентрация в головном и каудальном участках мозга выше, чем в центральной части.
Реакция ланцетника на свет выражается в форме отрицательного фототропизма. При искусственном освещении ланцетник закапывается в грунт. Рефлекторные ответы на световой стимул у ланцетника получа-ли только при условии морфофункциональной целостности головного участка мозга.
В головном участке имеются примитивные структуры - обонятельные ямки Келликера и инфундибулярный орган, которые рассматриваются в качестве прообразов переднего (обонятельного) и среднего мозга черепных хордовых. В передней стенке мозга имеется пигментное пятно, которое не обладает светочувствительностью, но функционально может быть аналогом эпифиза у рыб. Сенсорные системы ланцетника изучены плохо. Кроме светочувствительных глазков самого мозга, известны реснитчатые клетки кожи, чувствительные к гидродинамическим полям (прообраз боковой линии черепных рыб).
Особое место у ланцетника занимают первые две пары нервов. Они включают в себя чувствительные волокна, отходящие от предротовой области, пигментного пятна, жаберного аппарата и дыхательной мускулатуры.
Периферическая часть нервной системы ланцетника образована соматическими и вегетативными нервами и сплетениями. От нервного тяжа отходят 62-64 пары нервов, обеспечивающих сегментарный характер иннервации мышц. Эфферентные волокна нервов заканчиваются на мышцах особыми образованиями - концевыми конусами.
Внутренние органы иннервируются от дорзальных корешков. Трубчатые органы имеют нервные сплетения диффузного характера.
Следует отметить, что не только морфологически, но и функционально нервная система ланцетника уступает нервной системе черепных рыб (по чувствительности, скорости ответов и сложности нервных процессов). Однако ее можно принять за начальный этап цефализации нервной системы.
§12. НЕРВНАЯ СИСТЕМА КРУГЛОРОТЫХ
Промежуточное положение между примитивной нервной системой бесчерепных и развитой системой черепных рыб занимает нервная система круглоротых. У миног и миксин наблюдается дифференцирование головного и спинного мозга. Головной мозг у круглоротых достаточно примитивен (рис. 3.2).
В нем уже можно выделить передний, средний и задний отделы. Для круглоротых уже применимо понятие "нервный центр" как совокупность нейронов, отвечающих за определенную функцию. Однако для круглоротых все еще характерна высокая степень диффузности мозга.
В мозге круглоротых выявлены примитивные нисходящие проводящие пути, начало которых лежит в продолговатом мозге. Их основу составляют гигантские клетки, появившиеся у ланцетника.
У круглоротых можно найти хоть и примитивно устроенные, но уже настоящие сенсорные органы. Так, у миноги фоторецепция производится глазом, масса которого составляет более 20 % массы головного мозга. У миног и миксин хорошо развито обоняние, что подтверждается наличием специфических центральных нервных структур: габенулярного узла и обонятельных луковиц.
Для круглоротых применим термин "ганглионарный" тип строения вегетативного отдела периферической нервной системы.
У круглоротых интегрирующую функцию выполняют так называемый габенулярный узел (передний мозг), ретикулярная формация среднего мозга и подбугорная область. Сюда поступает вся афферентация. Габенулярный узел получает информацию от чувствительных нервных окончаний, отвечающих за химическую рецепцию. Средний мозг и гипоталамус соответственно осуществляют афферентный синтез световой рецепции и рецепции гидродинамических полей.
На этих участках головного мозга сходятся и все эфферентные соматические и вегетативные влияния.
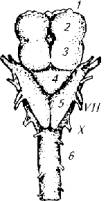
Рис. 3.2. Головной мозг миксины: 1 - обонятельный нерв; 2 - передний мозг; 3 - средний мозг: 4 -5- продолговатый мозг; 6 - спинной мозг; римскими цифрами обозначены головные нервы
Таким образом, на стадии круглоротых эволюция совершает существенный шаг вперед по пути цефализации нервной системы
Дифференциация отделов головного мозга вызвана скорее всего развитием сенсорных систем у рыб. Если у ланцетника отсутствует дифференциация мозга на головной и туловищный отделы при отсутствии сенсорных органов, то у круглоротых уже имеется сильно развитый передний мозг, обслуживающий афферентный анализ и синтез ольфакторной сенсорики. Средний мозг развит слабо, поскольку фоторецепция примитивна.
§13. НЕРВНАЯ СИСТЕМА ХРЯЩЕВЫХ РЫБ
У хрящевых рыб (акулы, скаты, химеры) есть три группы хорошо развитых сенсорных органов: химической рецепции, фоторецепции и органы акустико-латеральной системы. Соответственно происходит дифференциация головного мозга на три хорошо различимых отдела: передний (обонятельная луковица и обонятельная доля), средний (зрительные бугры) и задний (продолговатый мозг, мозжечок - отделы, собирающие афферентацию с органов акустико-латеральной системы). Степень развития того или иного отдела головного мозга отражает экологическую роль соответствующего сенсорного комплекса (рис. 3.3).
Определяющее значение в дифференциации головного мозга и всей центральной нервной системы имеют экологические факторы, что будет показано на примере костистых рыб. Здесь же отметим, что степень цефализации нервной системы у всех водных пойкилотермных животных (круглоротые, хрящевые рыбы, костистые рыбы, земноводные) примерно одинакова. Велика автономность спинного мозга у этих животных.
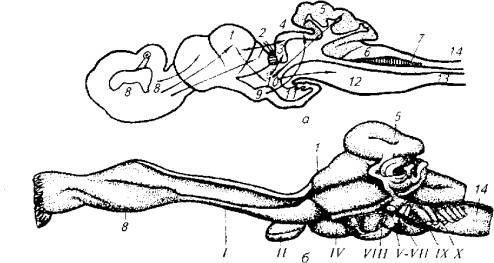
Рис. 3.3. Строение мозга хрящевых рыб:
а -акула; 6- химера; 1-передний мозг; 2- эпифиз; 3- габенулярный узел; 4-средний мозг; 5- мозжечок; 6, 7, 12- центры акустико-латеральной системы; 8 - обонятельная луковица; 9, 10,11 - структуры промежуточного мозга; 13 -- продолговатый мозг; 14- спинной мозг; римскими цифрами обозначены головные нервы
Существенный скачок в развитии нервной системы (ароморфоз, или идеоадаптация) появляется с выходом на сушу и приобретением животными гомойотермии.
§14. НЕРВНАЯ СИСТЕМА КОСТИСТЫХ РЫБ
У костистых рыб известно огромное количество особенностей (как правило, морфологического характера), которые ассоциируются как с видовой принадлежностью рыбы, так и с ее экологической нишей. Общими признаками для всех костистых рыб (как двоякодышащих Dipnoi, так и высших Teleostomi) являются одинаково хорошо развитая периферическая часть нервной системы, структурированный по функциям спинной мозг, сегментарный характер иннервации как опорно-двигательного аппарата, так и висцеро-вегетативных функций. Наиболее заметные различия между отдельными видами (или экологическими группами) рыб Osteichyes проявляются на уровне головного мозга.
Возникновение сложного сенсорного аппарата и морфофункциональное усложнение опорно-двигательного аппарата предопределили дальнейшую цефализацию нервной системы костистых рыб. Несмотря на разницу в дифференциации головного мозга в группе костистых рыб, для них характерны общие филогенетические приобретения: расширение связей между головным и туловищным отделом центральной нервной системы. Прежде всего это нисходящие пути, связывающие аппараты центральной регуляции движений со спинным мозгом: вестибулоспинальный тракт, церебеллоспинальный тракт и бульбоспинальный тракт.
При перерезке головного мозга выше продолговатого мозга, т. е. при сохранении бульбоспинального тракта, рыбы сохраняют способность к передвижению, но чувствительность животных и Реактивность к ряду раздражителей резко сокращаются, нарушайся координированность двигательных актов. Однако спинальная ры6a (после перерезки мозга между продолговатым и спинным отёлами) демонстрирует большую реактивность, что свидетельствует о дифференцированном тормозящем влиянии продолговатого Мозга на проявление спинальных рефлексов. Более того, первое время спинальные рыбы вообще утрачивают реактивность. У них развивается шоковое состояние, продолжающееся от нескольких минут до 1 часа.
Спинной мозг. Результаты исследований показали, что движения спинальных, медулярных (бульбарных) и мезэнцефальных рыб качественно различаются (рис. 3.4).
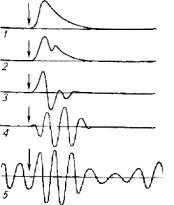
Рис. 3.4. Влияние уровня перерезки нервного тяжа золотой рыбки на характер движений ее хвостового стебля: 1-ниже 16-го позвонка; 2 - ниже 7-го позвонка; 3- спинальная рыба; 4- медулярная рыба; 5- мезэнцефальная рыба
У спинальных рыб отмечается рефлекторная активность всего тела, которая прекращается после стимуляции.
Для медулярных рыб характерны ритмические движения или всего туловища (вьюны, золотые рыбки), или только плавников (Sargus, Labrus, Serranus). Движения мезэнцефальных рыб еще сложнее. Причина различий заключается в наличии ганглиозных структур в продолговатом и среднем мозге, оказывающих нисходящее активизирующее влияние на спинной мозг. Клетки, лежащие в основании среднего мозга, выполняют функцию, аналогичную функциям красного ядра среднего мозга наземных позвоночных. Эти клетки (nucleus relicularis mesencephali) играют большую роль в регуляции координации движений рыб через распределение тонуса антагонистической мускулатуры. Данное явление характерно только для костистых рыб. У более примитивных рыб, например акул, спинной мозг имеет большую функциональную самостоятельность.
Расчленение головного и спинного мозга у хрящевых рыб не нарушает ритмическую активность спинного мозга при условии, что рыба находится во взвешенном положении. Если хрящевую спинальную рыбу положить на грунт, то она утрачивает ритмическую активность. Другими словами, даже у хрящевых рыб активность спинного мозга находится под влиянием головного мозга (по крайней мере, вестибулярного аппарата).
Большое влияние на рефлекторную двигательную активность спинного мозга оказывает периферическая афферентная импульсация, исходящая от внешних (кожных) и внутренних (мышечных, сухожильных) воспринимающих приборов.
Перерезка чувствительных волокон при сохранении эфферентных путей резко нарушает двигательную активность в пределах денервированного сегмента тела, что хорошо заметно на фоне соседних интактных сегментов. Полная деафферентация спинного мозга приводит к полной остановке ритмических движе-ний.
Интересно, что экспериментальная электрическая стимуляций спинного мозга деафферентированной рыбы не может заменить афферентный поток. Она вызывает лишь одиночные сокращения мыши туловища. Установлено, что биоэлектрическая активность
головного мозга поддерживается афферентным потоком, идущим кожи, мышц и сухожилий, постоянно как при раздражении чувствительного аппарата, так и в состоянии покоя. Роль афферентной стимуляции спинного мозга доказывают и такие опыты. При перерезке спинного мозга гольяна на уровне переднего края спинного плавника у рыбы исчезают спонтанные движения хвоста. Однако раздражение кожи рыбы восстанавливает ритмическую активность каудальной части спинного мозга и соответственно мышечную активность хвостового стебля. Афферентная стимуляция спинного мозга не является эволюционным приобретением костистых рыб. Она имеет глубокие эволюционные корни, поскольку хорошо развита у более древних животных - хрящевых рыб (рис. 3.5).
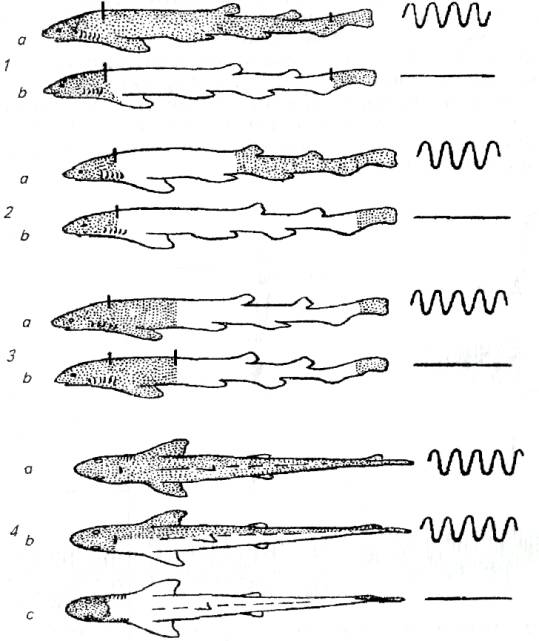
Рис. 3.5. Проявление ритмической двигательной активности акулы при различной деафферентации (светлое поле -зона деафферентации, серое поле-интактная зона)
Как следует из рис. 3.5, ритмическая двигательная активность зависит от топографии и площади деафферентации как у спинальных рыб (позиции 1, 2, 3), так и у рыб с интактным мозгом (позиция 4). Интересно, что односторонняя (даже тотальная) деафферентация не нарушает ритмической активности рыб при условии сохранения нисходящих путей из головного мозга. Последнее указывает на приоритетную роль головного мозга в поддержании ритмической активности спинного мозга и вторичность периферической афферентации,
При полной деафферентации и сохранении связи с головным мозгом в спинном мозге регистрируется высокая биоэлектрическая активность. Считают, что рефлекторные влияния головного мозга осуществляются по чувствительным путям сенсорных систем головного мозга. Биоэлектрическая активность спинного мозга при тотальной деафферентации последнего видоизменяется в ответ на звуковые и световые стимулы.
Доказаны влияние висцеральной афферентации на электрическую активность спинного мозга и его рефлекторное воздействие на вегетативные функции. При повышении или понижении давления газов в плавательном пузыре наблюдается усиление двигательной активности всех плавников рыбы, а также учащение дыхания и сердцебиения (рис. 3.6).
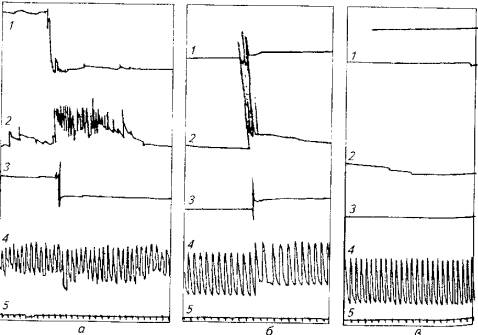
Рис. 3.6. Афферентные влияния плавательного пузыря на движение хвоста (1), плавников (2), частоту дыхания (3), сердечный ритм (4, 5)
Афферентный поток меняется как при изменении давления газов в плавательном пузыре (см. рис. 3.6, а), так и при раздражении вагосимпатического нерва плавательного пузыря (см. рис. 3,6, б}. Нервный характер описанного явления доказывает рис. 3.6, в, на котором отсутствует возбуждающий эффект раздражения плавательного пузыря после перерезки вагосимпатического нерва (X. С. Коштоянц, 1957).
Помимо соматических рефлексов спинной мозг рыб обеспечивает регуляцию практически всех висцеральных рефлексов. В спинном мозге расположены нейроны, которые регулируют функции всех внутренних органов по принципу безусловных рефлексов.
Таким образом, рефлекторная активность спинного мозга проявляется как результат взаимодействия его нейронов со структурами головного мозга при постоянном активизирующем афферентном влиянии со стороны опорно-двигательного аппарата и висцеральных органов.
Условно-рефлекторная активность спинного мозга рыб отвергается большинством ученых. Считают, что временные связи в спинном мозге рыб не образуются.
Головной мозг. Строение головного мозга костистых рыб прежде всего связано с этологическими особенностями этих животных. Анализ анатомии головного мозга и развития сенсорных систем позволяет утверждать, что развитие сенсорных систем привело к цефализации нервной системы в целом и дифференциации головного мозга в частности.
В табл. 3.1 приведены данные о связи между степенью развития головного мозга и глаз. Головной мозг занимает в среднем около 0,15 % массы тела костистых рыб. Однако межвидовые различия довольно велики: от 0,6 % у карася до 0,03 % у угря. Большое соотношение массы глаз и массы головного мозга приходится на хищных рыб (щука, форель, жерех, голавль).
Соотношение масса глаз: масса мозга у костистых рыб в 4 раза больше по сравнению с хрящевыми рыбами. Это согласуется с этологическими наблюдениями, которые свидетельствуют о том, что зрительный анализатор у костистых рыб более развит и играет большую роль в формировании мотиваций поведения.
Продолговатый мозг. Продолговатый мозг занимает большую часть заднего мозга. Он расположен на границе спинного и головного мозга. Поэтому прежде всего на него ложится проводниковая функция. В его состав входят как нисходящие, так и восходящие пути, соединяющие спинной мозг с разными отделами головного мозга. Кроме того, в составе продолговатого мозга имеются гигантские клетки, образующие с ганглиозными клетками сетчатую Формацию, которая формирует собственные проводящие пути продолговатого мозга.
В продолговатом мозге рыб лежат ядра шести нервов: с V по X.
Ядра черепно-мозговых нервов у разных рыб имеют разную степень развития: от невыраженного ядра VI нерва (отводящего) до гипертрофированных вагусных долей X нерва (блуждающий) у карповых рыб.
Дата добавления: 2014-12-11 | Просмотры: 1913 | Нарушение авторских прав
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 34 | 35 |
|