|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология
|
Характеристика красных и белых мышц
| Красные мышцы
| Белые мышцы
| | Обеспечивают медленные движения рыб
| Обеспечивают быстрые движения рыб
| | Преобладание анаэробных процессов
| Преобладание аэробных процессов
| | Богаты гликогеном
| Гликогена в 2 раза меньше, чем в красных мышцах
| | Функциональные нагрузки влияют на уровень гликогена незначительно
| Резкое падение уровня гликогена при быстром плавании
| | Богаты жиром
| Небольшое содержание жира (20 % содержания жира в красных мышцах)
| | Богаты миоглобином
| Миоглобин отсутствует
|
Красные мышцы по массе уступают белым. Их количество у большинства рыб составляет несколько процентов от массы белой мускулатуры. И лишь у активных пловцов, находящихся в постоянном движении - тунцов, ставриды, макрели, сельди -доля красной мускулатуры достигает 20 %.
Красная мускулатура находится над белой мускулатурой, непосредственно под кожей, в основном в латеральной области хвостового стебля и туловища рыбы.
Лишь у видов, использующих для движения грудные плавники, красная мускулатура локализована в непосредственной близости от плавников и жаберных крышек. У активных пловцов (например, тунца) красные мышцы пронизывают всю толщу белых мышц и достигают позвоночного столба (рис. 5.8).
Темный цвет мускулатуре придают железосодержащие белки. Мышечные волокна содержат большое количество миоглобина и цитохромов. Кроме того, темная мускулатура имеет обильную васкуляризацию. В состоянии покоя кровоток в красных мышцах в 3 раза превышает таковой в белых мышцах. При возбуждении приток крови к темным мышцам многократно возрастает.
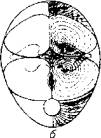
Рис. 5.8. Топография красной мускулатуры на поперечном разрезе тела тунца
Функционально красная мускулатура также выделяется. Она обеспечивает мед. ленную, но продолжительную работу. Красные мышцы меньше утомляются и быстрее восстанавливаются при интенсивных нагрузках. Именно в такого рода мышечной деятельности испытывают повышенную потребность активные пелагические рыбы - тунцы и макрели, покрывающие за сутки расстояния в сотни и тысячи километров.
В качестве источника энергии красные мышцы используют жирные кислоты и кетоновые тела, что возможно лишь при наличии кислорода. В красных мышцах содержится мало гликогена и много ферментов, необходимых для аэробных реакций. Даже при предельном напряжении в красных мышцах активно протекают аэробные процессы. Поэтому так велика потребность красных мышц в кислороде и так развита в них сеть капиллярных сосудов.
Мышечные волокна красной мускулатуры иннервируются медленными нервными волокнами, т. е. волокнами малого диаметра. Это частично объясняет причины большого латентного периода красных мышц. Однако на каждом мышечном волокне имеется несколько концевых пластинок, гарантирующих точность выполнения команды из центральной нервной системы.
Светлые мышцы у рыб составляют основную массу активной части опорно-двигательного аппарата. Они отличаются более коротким латентным периодом (по сравнению с темными мышцами), большой скоростью сокращения и расслабления. Как для светлых, так и для красных мышц характерны тетанические сокращения, что определено природой электрического импульса, поступающего по аксону мотонейрона, - спайк с частотой 30- 50 Гц. Иннервация светлых мышц осуществляется за счет быстрых (миелинизированных) нервных волокон большого диаметра. Каждое мышечное волокно у костистых рыб имеет несколько нервно-мышечных синапсов.
Кровоснабжение светлых мышц не такое интенсивное, как красных. В качестве источника энергии светлые мышцы используют глюкозу. Тот факт, что в возбужденных светлых мышцах всегда присутствует молочная кислота, свидетельствует о приоритете гликолиза в процессе мышечного сокращения.
Приверженность светлых мышц к анаэробному использованию энергии глюкозы подчеркивает и высокая активность ферментов анаэробного цикла в светлых мышцах рыб. Известно, что гликолиз - неэф-фективный путь использования энергии по сравнению с окислительным фосфорилированием. В данном случае природа приносит в жертву энергетическую экономию для получения выигрыша в скорости мышечного сокращения.
Клетка мышечной ткани (мышечное волокно) имеет специфическое строение. Снаружи она покрыта особой мембранной оболочкой - сарколеммой, а ее протоплазма (саркоплазма) наряду с традиционными цитозольными структурами включает в себя особые органеллы - миофибриллы,
Миофибрилла является структурным элементом мышечного волокна, обеспечивающим укорочение (сокращение) (рис. 5.9).
При световом микроскопировании миофибрилл видна их поперечная исчерченность из-за разницы в светопреломлении входящих в их состав белков - актина и миозина. В состав миофибрилл входят толстые и тонкие протофибриллы. Тонкие актиновые протофибриллы образуют так называемые изотропные диски (I-диски). В средней части изотропного диска расположена Z-пластина.
Более толстые миозиновые протофибриллы и заходящие своими концами актиновые нити образуют анизотропные диски (А-диски). В средней части анизотропных дисков тонких нитей нет, поэтому эту зону выделяют в самостоятельную Н-зону. В области взаимного перекрытия в пределах А-диска актиновые и миозиновые нити соединены при помощи особых мостиков. Кроме того, актиновые нити закреплены на поперечных Z-пластинах (рис. 5.10).
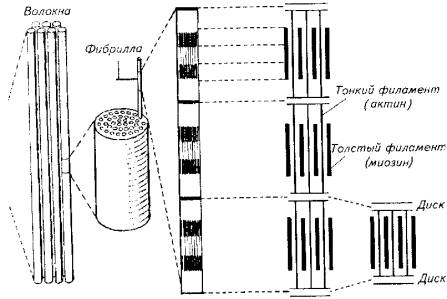
Рис. 5.9. Микроструктура скелетной мышцы
Исследования позволили выявить в составе тонких актиновых нитей и элементы другого белка - миозина (в виде тропомиозина).
Участок миофибриллы между двумя Z-пластинами называют саркомером.
Количество и соотношение толстых и тонких протофибрилл колеблется не только у разных видов рыб, но и в разных мышцах у одной особи. Быстрые мышцы имеют большее количество нитей и меньше саркоплазмы. В медленных мышцах больше саркоплазмы.
Помимо нитевидных структур - миофибрилл и протофибрилл, в составе мышечного волокна скелетных мышц выделяется хорошо развитая тубулярная система - сеть каналов и трубочек. Подсчитано, что общая поверхность мембран тубулярной системы в 10 раз превышает площадь наружной мембраны мышечного волокна. Сарколемма образует разветвленные выпячивания, проникающие в область Z-пластин. Их называют Т-канальцами. К Т-канальцам прилегает другая тубулярная система, которая относится к саркоплазматическому ретикулуму. Она образует сложную ячеистую сеть вокруг саркомера (рис. 5.11).
Эта мембранная сеть обладает очень высокой ферментативной активностью. Она же выступает в роли депо ионизированного кальция, без которого невозможно мышечное сокращение.
В медленных мышцах объем Т-канальцев значительно меньше по сравнению с их объемом в быстрых мышцах.
Потенциал действия, возникающий на сарколемме при возбуждении мышцы, передается по Т-канальцам, а затем по каналам саркоплазматического ретикулума достигает непосредственно протофибриллярного аппарата.
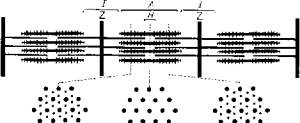
Рис. 5.10. Взаимное расположение актиновых и миозиновых протофибрилл
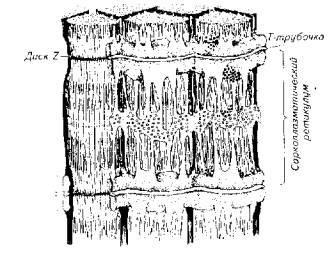
Рис. 5.11. Схема, иллюстрирующая расположение фибриллярных элементов и тубулярной сети мышечного волокна
В целом скорость сокращения мышцы зависит от длины саркомера, количества протофибрилл и характера их расположения в пределах саркомера, количества митохондрий и энергетического материала, наличия и количества миоглобина в саркоплазме.
§23. ГЛАДКИЕ МЫШЦЫ
Гладкие мышцы образуют стенки внутренних трубчатых органов. Микроструктура этого типа мышц не так однородна, как микроструктура скелетной мускулатуры. У них нет поперечной исчерченности, так как нет упорядочение расположенных толстых и тонких протофибрилл. Во многих висцеральных мышцах толстых протофибрилл вообще нет. Однако при химическом анализе мыши в них обязательно обнаруживается белок миозин. Считают, что в последнем случае сокращение мышечного волокна происходит за счет полимеризации молекул миозина.
В составе некоторых гладких мышц обнаружены очень толстые Темные протофибриллы сложного строения. Их основу составляет белок тропомиозин, а на поверхности находится миозин.
Помимо специфики фибриллярного аппарата гладкие мышцы отличаются от поперечнополосатых еще и слаборазвитой тубулярной системой. Так, у гладких мышц нет Т-трубочек. Саркоплазматическая система слабо развита (в некоторых мышцах вообще отсутствует) и представлена своеобразными пузырьками, Протофибриллярный аппарат имеет непосредственные зоны соприкосновения с сарколеммой.
Волокна гладких мышц могут быть как длинными, так и короткими. У рыб и те и другие образуют контакты друг с другом. Не все волокна в гладкой мышце имеют контакты с нервным окончанием. Поэтому двигательная единица гладких мыши имеет специфическое строение. Она включает нервное окончание вегетативного нерва, концевую пластинку и группу из 5-10 мышечных клеток, которые имеют контакты между собой. Естественно, что такая единица реагирует на нервную стимуляцию очень медленно.
Гладкие мышцы способны к спонтанному возбуждению. Большинство висцеральных органов имеет интрамуральные нервные сплетения, которые обеспечивают высокую степень автономности трубчатых органов.
Гладкие мышцы в отличие от скелетных не чувствительны к электрической стимуляции и очень чувствительны к механическому раздражению (например, растяжению). Отличаются гладкие мышцы и своей высокой реактивностью по отношению к химическим раздражителям (ацетилхолин, норадреналин. окситоцин, соли калия, кальция, магния, иногда желчь, соляная кислота), что не свойственно поперечнополосатым мышцам.
Сокращения гладких мышц носят характер медленных и продолжительных, что и требуется для обеспечения перемешивания и продвижения химуса по пищеварительной трубке, выделения желчи, мочи, извержения половых продуктов при икрометании и т. д.
§24. МЕХАНИЗМ МЫШЕЧНОГО СОКРАЩЕНИЯ
События, развивающиеся на микроскопическом (субклеточном) уровне при возбуждении мышцы, выглядят следующим образом. Потенциал действия, возникающий в результате нервной или иной стимуляции на мембране мышечного волокна, в скелетных мышцах по Т-каналам достигает мембран цитоплазматического ретикулума. Мембрана последнего высвобождает ионизированный кальций, который, в свою очередь, активизирует процесс взаимодействия актиновых и миозиновых протофибрилл.
В гладких мышцах, лишенных тубулярной системы, высвобождение Са++ происходит непосредственно в самой сарколемме. А так как сарколемма напрямую контактирует с актомиозиновым комплексом гладких мышц, то активизация сокращения гладких мышц также происходит под влиянием ионизированного кальция.
Интимный механизм взаимодействия актиновых и миозиновых структур до конца не расшифрован. Однако ясно, что в присутствии АТФ в поперечнополосатых мышцах при выбросе Са++ начинается скольжение тонких и толстых протофибрилл относительно друг друга (рис. 5.12). Считается, что непосредственной причиной этого движения протофибрилл является активизация актомиозиновых мостиков, которые либо изменяет угол наклона, либо подвергаются торсионному скручиванию (рис. 5.12).
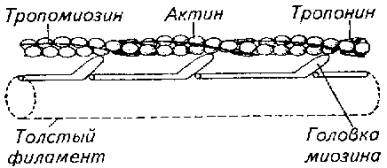
Рис. 5.12. Модель взаимодействия актиновой и миозиновой нитей
Установлена АТФ-азная зависимость конформационных изменений актомиозиновых мостиков. В отсутствие АТФ эти изменения невозможны, т. е. процесс скольжения протофибрилл не происходит.
Следует подчеркнуть исключительно важную роль ионов кальция в этом процессе, который не только влияет на конформационные изменения актомиозина, но и является активатором АТФ-азы. Более того, процесс расслабления мышцы зависит от Са++. Актиновые и миозиновые нити не возвращаются в исходное положение до тех пор, пока не произойдет обратное поглощение ионов кальция мембраной цитоплазматического ретикулума или мембраной сарколеммы (в гладких мышцах), причем это обратное движение кальция зависит от активности АТФ-азы соответствующего мембранного комплекса. Поэтому не только сокращение, но и расслабление мышцы протекает с затратами энергии.
Эксперименты показали, что АТФ-азную активность проявляет сам белок миозин. Он же при наличии АТФ способен соединяться с актином. Причем АТФ-азная часть миозина активируется актином. Однако белок тропонин в составе актиновой нити блокирует эту активность. И только выброс Са++ снимает тормозящее действие тропонина. Поэтому тропонин-тропомиозиновую систему рассматривают как предохранительный механизм взаимодействия актиновых и миозиновых нитей в состоянии физиологического покоя.
Механизм укорочения гладкомышечного волокна менее понятен. Распространена точка зрения, согласно которой укорочение гладкомышечного волокна есть результат конформационного изменения актомиозинового комплекса. Белок актомиозин выделен из гладких мышц многих животных, включая рыб. По своим свойствам он близок к миозину млекопитающих. Как актиновая, так и миозиновая молекула имеют фибриллярную спиралевидную часть. Активизация актомиозина приводит к изменению конформации молекулы белка с соответствующим изменением его пространственного внутрицеллюлярного положения. Это и приводит к общему укорочению мышечного волокна, так как актомиозиновые структуры морфологически связаны с сарколеммой гладкомышечного волокна.
Нервная регуляция гладкомышечной активности у рыб довольно специфична и заслуживает специального обсуждения. Влияние нервной системы на гладкие мышцы может быть как возбуждающим, так и тормозящим. Характер нервной стимуляции гладких мышц существенно зависит от гормонального статуса рыб и функционального состояния самого висцерального органа (например, степени наполнения желудка, мочеточников, стадии созревания половых клеток в ястыке и их наличия в матке или яйцеводе и т. д.).
Гладкая мускулатура иннервируется симпатическим и парасимпатическим отделами нервной системы. Кроме того, многие висцеральные органы (например, желудочно-кишечный тракт крупные сосуды) обладают собственной интрамуральной нервной системой в виде нервных сплетений разной степени сложности.
Как правило, симпатические нервные волокна в качестве медиаторов выделяют норадреналин, а парасимпатические волокна - соответственно ацетилхолин. Однако исследования показали, что в составе парасимпатической системы есть адренергические, а в составе симпатической системы -холинергические волокна. Поэтому у рыб симпатическая и парасимпатическая нервные системы являются смешанными по функциональным признакам. У ацетилхолина имеется два типа рецепторов - мускариновые и никотиновые. Для норадреналина описано также два функциональных типа рецепторов, называемых а- и р-рецепторами.
Стимуляция вагуса как представителя парасимпатической части нервной системы у рыб вызывает различные эффекты не только по отношению к различным физиологическим системам, но и в пределах одной системы. Так, у форели и других желудочных рыб вагус тормозит работу желудка. Возбуждающему эффекту желудок подвергается при стимуляции чревного нерва.
Далее желудка (кишечник) влияние симпатической и парасимпатической систем изменяется на прямо противоположное. Адренергические волокна и адреналин оказывают тормозящее действие, а холинергические волокна и ацетилхолин стимулируют перистальтику кишечника. Интересно и то, что ацетилхолин возбуждает все отделы кишечника у рыб. Адреналин же возбуждает только желудок и оказывает тормозящее действие на весь кишечник.
У некоторых рыб пищеварительная трубка содержит элементы поперечнополосатой мускулатуры. Например, у линя кишка покрыта своеобразным чехлом подобных мышц. Поэтому его кишечник одинаково отвечает как на электрическую, так и на гуморальную стимуляцию.
Кроме плавания и висцеральных функций мышцы обеспечивают и некоторые довольно специфичные движения рыб. Так, таиландский ходячий сом в составе грудных плавников имеет шипы-ходули, которыми он пользуется для переползания по суше из одного водоема в другой. Наличие таких ходуль и приспособленность к дыханию атмосферным воздухом (имеет хорошо развитую систему наджаберных полостей) позволяют этой рыбе преодолевать в период засухи довольно большие расстояния в поисках
Правда, при движении по суше (точнее сказать грязи, так как рыба для передвижения все-таки выбирает увлажненные места) сом использует не только грудные плавники-ходули, но и при вытянутом теле, извиваясь подобно змее, облегчает работу мышц плавников. Эта особенность таиландского сома позволила ему освоить большие территории на юге США, куда он случайно попал из Азии.
Морской петух тригла (рис. 5.13) также ползает по дну, но при этом опирается сразу на три луча грудных плавников (отсюда и его название "тригла"). Интересно, что эти ходильные лучи у петуха обособлены от других и способны сгибаться. Кроме движения по дну видоизмененные лучи грудных плавников обеспечивают этой рыбе тактильную рецепцию.
В других случаях рыбы используют грудные плавники для планирования в воздухе. Так, летучие рыбы при испуге за счет броскового движения туловищных мышц, мышц хвостового стебля и интенсивной работы нижней лопастью хвостового плавника выскакивают из воды и пролетают в воздухе расстояния, по-зволяющие им избавиться от преследователей (рис. 5.14).
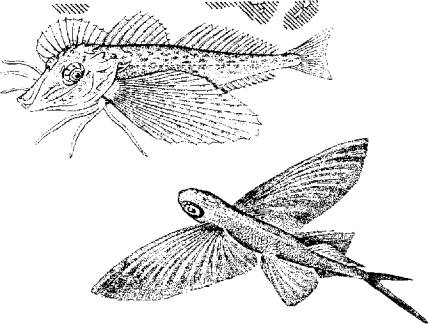
Рис. 5.13. Морской петух использует лучи грудных плавников для "хождения" по дну. Рис. 5.14. Полет пальцекрыла
На поверхности воды летучая рыба достаточно продолжительно работает хвостом, развивая большую тягу, позволяющую ей преодолеть силу притяжения. Скорость полета этих мелких рыб превышает скорость движения преследователей (тунцы, меч-рыба), а пролетаемые ими расстояния достигают нескольких сотен метров.
Другие виды рыб, например пальцекрыл, могут не только парить, но и выполнять сложные маневры в воздухе. Пальцекрыл поднимается к поверхности воды и скользит по ней со скоростью 18м/с. Такую высокую скорость рыба приобретает благодаря зигзагообразным движениям хвостового плавника с гипертрофированной нижней лопастью (рис. 5.15).
Сильный удар хвоста поднимает рыбу в воздух на высоту 5-7 м. Пальцекрыл пролетает в воздухе до 200м, используя при этом и воздушные потоки. Рыба способна при необходимости изменить направление полета за счет движений хвостового плавника. У нее также отмечены колебательные движения грудных плавников.
Скорость полета пальцекрыла сопоставима со скоростью движения современных морских судов и нередко достигает 60- 70 км/ч,
Интересен механизм движений, приводящий в "боевое" положение колючки плавников таких рыб, как ерши, окуни, колюшки, сомы. Острые и крепкие колючки плавников этих рыб защищают их от хищников.
В случае опасности плавники расправляются, а колючки поднимаются перпендикулярно телу. При этом несколько увеличиваются границы тела рыбы. Проглотить или схватить такую рыбу может не каждый хищник, так как надавливание на колючки не приводит к их складыванию. Следовательно, хищник при захвате неизбежно травмирует ротовую полость. Вероятнее всего, в следующий раз хищник откажется от нападения на такую рыбу.

Рис. 5.15. След на поверхности воды, оставляемый летучей рыбой при взлете
Кроме того, расправленные плавники и торчащие колючки ложно увеличивают размеры рыбы, что также повышает ее шансы на выживание.
Важно отметить, что колюшкам и сомам не приходится прибегать к мышечной силе, так как колючки в "боевом" положений
удерживаются не мышцами, а при помощи особого механизма. (Рис. 5.16.)
B механизме фиксации колючки заложен принцип замка, который используется и другими животными (коленный сустав лошади, суставы нижних конечностей вороны и др.). Чем сильнее хищник давит на колючку, тем прочнее колючка фиксируется в замке, т е. прочность замка возрастает за счет использования энергии хищника, а не потенциальной жертвы.
Опорой для колючки и ее замка служит особое образование из нескольких костей и мышц, формирующих шарнир (см. рис.5.16,///)
Шарнир запирается при помощи V-образной косточки, которая имеет прочные связки с колючкой и скелетом. При возбуждении рыбы мышцы 5 и 6 сокращаются, колючка поднимается, а кость 3 блокирует шарнир. В таком положении давление на колючку делает фиксацию шарнира только жестче.
Для опускания колючки необходимо, чтобы расслабились мышцы 5, 6 и 7, но сократилась мышца 8. На удержание колючки в поднятом состоянии расходуется минимум мышечных усилий, а для приведения в горизонтальное положение требуется сокращение всего одной мышцы 8.
Рыбы-брызгуны используют мышечный аппарат глотки, рта и жабр для выплевывания струи воды (рис. 5.17).
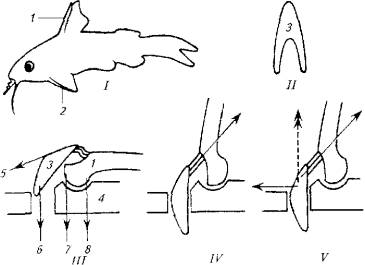
Рис. 5.16. Схема фиксации плавниковых колючек:
I- общий вия рыбы; III- IV- разные положения сочленения костей; 1 - колючка спинного плавника; 2- колючка грудного плавника; 3- опорный скелет; 4, II-V-образная косточка; 5-8- места прикрепления мышц

Рис. 5.17. Рыба-брызгун: охота на насекомых при помощи струи воды
Рыба-брызгун достигла большого совершенства в прицельной стрельбе по насекомым, При длине собственного тела не более 20 см рыба выбрасывает мощную струю воды, которая сбивает насекомых на расстоянии до 1,5 м от поверхности воды. Впечатляет и высокая результативность такой охоты. Промахи у этой рыбы - крайне редкое явление.
Все это разнообразие движений, наблюдаемое в классе рыб, обеспечивает сложная система опорно-двигательного аппарата и гладкомышечных образований, объединенных в единое целое при помощи нервной системы. Движения рыбы, в какой бы форме они ни проявлялись, осуществляют оперативное реагирование организма на изменения среды обитания с целью избежания опасности или адаптации к новым условиям.
§25. СКОРОСТЬ ДВИЖЕНИЯ
Скорость плавания рыбы в воде пропорциональна частоте и амплитуде колебаний тела и хвоста и укладывается в рамки определенных математических уравнений. Например, для карася, ельца и форели предельная скорость плавания
v=(1/4)[L(3f-4)],
где L - длина тела; f- частота колебаний тела (хвоста).
Несмотря на то что вода является более плотной средой по сравнению с воздухом, рыбам удается достичь очень высоких скоростей (км/ч):
Рыба-меч 130
Голубой марлин 90
Голубой тунец 80
Терпуг 70
Голубая акула 60
Атлантический лосось 40
Естественно, что для обеспечения таких скоростей природа наделила рыб рядом морфологических, гидродинамических и метаболических приспособлений.
Поскольку предельные абсолютные скорости рыб зависят от их чиненных размеров, специалисты предлагают использовать для сопоставления скоростных возможностей рыб относительный показатель - коэффициент скорости:
Этот коэффициент характеризует скорость рыбы, равную числу ее корпусов (длин) в секунду. Тогда все виды рыб можно классифицировать, как минимум, шестью категориями (табл. 5.4). Понятно, что рыбы с одинаковыми максимальными скоростями движения, но с различной длиной тела согласно этой классификации могут относиться к разным категориям.
Дата добавления: 2014-12-11 | Просмотры: 1636 | Нарушение авторских прав
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 34 | 35 |
|