|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология
|
Глава 7. ОСОБЕННОСТИ КРОВООБРАЩЕНИЯ РЫБ
Кровь выполняет многочисленные функции только тогда, когда движется по сосудам. Обмен веществ между кровью и другими тканями организма происходит в капиллярной сети. Отличаясь большой протяженностью и разветвленностью, она оказывает большое сопротивление току крови. Давление, необходимое для преодоления сопротивления сосудов, создается в основном сердцем,
Строение сердца рыб проще, чем высших позвоночных. Производительность сердца у рыб как нагнетательного насоса значительно ниже, чем у наземных животных. Тем не менее оно справляется со своими задачами. Водная среда создает благоприятные условия для работы сердца. Если у наземных животных значительная часть работы сердца затрачивается на преодоление сил гравитации, вертикальные перемещения крови, то у рыб плотная водная среда существенно нивелирует гравитационные влияния. Вытянутое в горизонтальном направлении тело, небольшой объем крови, наличие только одного крута кровообращения дополнительно облегчают функции сердца у рыб.
§30. СТРОЕНИЕ СЕРДЦА
Сердце у рыб небольшое, составляющее примерно 0,1% массы тела. Из этого правила, конечно, есть исключения. Например, у летучих рыб масса сердца достигает 2,5 % массы тела.
Для всех рыб характерно двухкамерное сердце. Вместе с тем существуют видовые различия в строении этого органа. В обобщенном виде можно представить две схемы строения сердца в классе рыб. И в первом, и во втором случае выделяют 4 полости: венозный синус, предсердие, желудочек и образование, отдаленно напоминающее дугу аорты у теплокровных, - артериальную луковицу у костистых и артериальный конус у пластинчатожаберных (рис, 7.1).
Принципиальное различие этих схем заключено в морфофункциональных особенностях желудочков и артериальных образований.
У костистых артериальная луковица представлена фиброзной тканью с губчатым строением внутреннего слоя, но без клапанов.
У пластинчатожаберных артериальный конус помимо фиброзной ткани содержит и типичную сердечную мышечную ткань, поэтому обладает сократимостью. Конус имеет систему клапанов, облегчающих одностороннее продвижение крови через сердце.
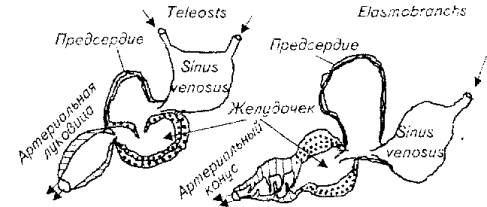
Рис. 7.1. Схема строения сердца рыб
В желудочке сердца рыб обнаружены различия в структуре миокарда. Принято считать, что миокард рыб специфичен и представлен однородной сердечной тканью, равномерно пронизанной трабекулами и капиллярами. Диаметр мышечных волокон у рыб меньше, чем у теплокровных, и составляет 6-7 мкм, что вдвое меньше по сравнению, например, с миокардом собаки. Такой миокард называют губчатым.
Сообщения о васкуляризации миокарда рыб довольно запутанны. Миокард снабжается венозной кровью из трабекулярных полостей, которые, в свою очередь, заполняются кровью из желудочка через сосуды Тибезия (Thebesian vessels). В классическом понимании у рыб нет коронарного кровообращения. По крайней мере, медики-кардиологи придерживаются такой точки зрения. Однако в литературе по ихтиологии термин "коронарное кровообращение рыб" встречается часто.
В последние годы исследователи обнаружили много вариаций васкуляризации миокарда. Например, С. Agnisola et. al (1994) сообщает о наличии двуслойного миокарда у форели и электрического ската. Со стороны эндокарда лежит губчатый слой, а над ним слой миокардиальных волокон с компактным упорядоченным расположением.
Исследования показали, что губчатый слой миокарда обеспечивается венозной кровью из трабекулярных лакун, а компактный слой получает артериальную кровь по гипобронхиальным артериям второй пары жаберных дут. У elasmobranchs коронарное кровообращение отличается тем, что артериальная кровь из гипобронхиальных артерий доходит до губчатого слоя по хорошо развитой системе капилляров и попадает в полость желудочка по сосудам Тибезия.
Еще одно существенное различие костистых и пластинчатожаберных заключается в морфологии перикарда.
У костистых перикард напоминает таковой наземных животных. Он представлен тонкой оболочкой.
У пластинчатожаберных перикард образован хрящевой тканью поэтому он представляет собой как бы жесткую, но упругую капсулу. В последнем случае в период диастолы в перикардиальном пространстве создается некоторое разрежение, что облегчает кровенаполнение венозного синуса и предсердия без дополнительных затрат энергии.
§31. ЭЛЕКТРИЧЕСКИЕ СВОЙСТВА СЕРДЦА
Строение миоцитов сердечной мышцы рыб сходно с таковым высших позвоночных. Поэтому и электрические свойства сердца похожи. Потенциал покоя миоцитов у костистых и пластинчатожаберных составляет 70 мВ, у миксин - 50 мВ. На пике потенциала действия регистрируется изменение знака и величины потенциала с минус 50 мВ до плюс 15 мВ. Деполяризация мембраны миоцита приводит к возбуждению натрий-кальциевых каналов. Сначала ионы натрия, а затем ионы кальция устремляются внутрь клетки миоцита. Этот процесс сопровождается образованием растянутого плато, а функционально фиксируется абсолютная рефрактерность сердечной мышцы. Эта фаза у рыб значительно продолжительнее - около 0,15 с.
Следующая за этим активизация калиевых каналов и выход ионов калия из клетки обеспечивают быструю реполяризацию мембраны миоцита. В свою очередь, реполяризация мембраны закрывает калиевые и открывает натриевые каналы. В итоге потенциал клеточной мембраны возвращается к исходному уровню минус 50 мВ.
Миоциты сердца рыбы, способные к генерации потенциала, локализованы в определенных участках сердца, которые совокупно объединены в "проводящую систему сердца". Как и у высших позвоночных, у рыб инициирование сердечной систолы происходит в синатриальном узле.
В отличие от других позвоночных у рыб роль пейсмейкеров выполняют все структуры проводящей системы, которая у костистых включает в себя центр ушкового канала, узел в атриовентрикулярной перегородке, от которого к типичным кардиоцитам желудочка тянутся клетки Пуркинье.
Скорость проведения возбуждения по проводящей системе сердца у рыб ниже, чем у млекопитающих, причем в разных участках сердца она неодинакова. Максимальная скорость распространения потенциала зарегистрирована в структурах желудочка.
Электрокардиограмма рыб напоминает электрокардиограмму человека в отведениях V3 и V4 (рис. 7.2). Однако техника наложений отведений для рыбы не разработана так подробно, как для наземных позвоночных животных.

Рис. 7.2. Электрокардиограмма рыбы
У форели и угря на электрокардиограмме хорошо видны зубцы Р, Q, R, S и Т. Только зубец S выглядит гипертрофированным, а зубец Q неожиданно имеет положительную направленность, у пластинчатожаберных в дополнение к пяти классическим зубцам на электрокардиограмме выявлены зубцы Bd между зубцами S и Т, а также зубец Вr между зубцами Г и.Р. На электрокардиограмме угря зубцу Р предшествует зубец V. Этиология зубцов такова:
зубец Р соответствует возбуждению ушкового канала и сокращению венозного синуса и предсердия;
комплекс QRS характеризует возбуждение атриовентрикулярного узла и систолу желудочка;
зубец Т возникает в ответ на реполяризацию клеточных мембран сердечного желудочка.
§32. РАБОТА СЕРДЦА
Сердце рыб работает ритмично. Частота сердечных сокращений у рыб зависит от многих факторов.
Частота сердечных сокращений (ударов в минуту) у карпа при 20 њС
Личинка
Молодь массой 0,02 г 80
Сеголетки массой 25 г 40
Двухлетки массой 500 г 30
В опытах in vitro {изолированное перфузированное сердце) частота сердечных сокращений у радужной форели и электрического ската составила 20-40 ударов в минуту.
Из множества факторов наиболее выраженное влияние на частоту сердечных сокращений оказывает температура среды обитания. Методом телеметрии на морском окуне и камбале была выявлена следующая зависимость (табл. 7.1).
7.1. Зависимость частоты сердечных сокращений от температуры воды
| Температура, њС
| Частота сердечных сокращений, ударов в минуту
| Температура, њС
| Частота сердечных сокращений, ударов в мнуту
| |
|
| 11,5
|
| |
|
|
|
| |
|
|
|
|
Установлена видовая чувствительность рыб к перепадам температуры. Так, у камбалы при повышении температуры воды с g до 12 њС частота сердечных сокращений возрастает в 2 раза (с 24 до 50 ударов в минуту), у окуня - только с 30 до 36 ударов в минуту.
Регуляция сердечных сокращений осуществляется при помощи центральной нервной системы, а также внутрисердечных механизмов. Как и у теплокровных, у рыб в опытах in vivo при повышении температуры притекающей к сердцу крови наблюдалась тахикардия. Понижение температуры притекающей к сердцу крови вызывало брадикардию. Ваготомия снижала уровень тахикардии.
Хронотропным действием обладают и многие гуморальные факторы. Положительный хронотропный эффект получали при введении атропина, адреналина, эптатретина. Отрицательную хронотропию вызывали ацетилхолин, эфедрин, кокаин.
Интересно, что один и тот же гуморальный агент при различной температуре окружающей среды может оказывать прямо противоположное воздействие на сердце рыб. Так, на изолированном сердце форели при низких температурах (6њС) эпинефрин вызывает положительный хронотропный эффект, а на фоне повышенных температур (15њС) перфузирующей жидкости - отрицательный хронотропный эффект.
Сердечный выброс крови у рыб оценивается в 15-30 мл/кг в минуту. Линейная скорость крови в брюшной аорте составляет 8- 20 см/с. In vitro на форели установлена зависимость сердечного выброса от давления перфузирующей жидкости и содержания в ней кислорода. Однако в тех же условиях у электрического ската минутный объем не изменялся.
В состав перфузата исследователи включают более десятка компонентов.
Состав перфузата для сердца форели (г/л)
Хлорид натрия 7,25
Хлорид калия 0,23
Фторид кальция 0,23
Сульфат магния (кристаллический) 0,23
Фосфат натрия однозамещенный (кристаллический) 0,016
Фосфат натрия двузамещенный (кристаллический) 0,41
Глюкоза 1,0
Поливинил пиррол идол (PVP) коллоидный 10,0
Примечания:
I. Раствор насыщается газовой смесью из 99,5 % кислорода, 0,5 % углекислого газа (диоксида углерода) или смесью воздуха (99 5%) с углекислым газом (0,5 %).
2.рН перфузата доводят до 7,9 при температуре 10 њС, используя бикарбонат натрия.
Состав перфузата для сердца электрического ската (г/л)
Хлорид натрия 16,36
Хлорид калия 0,45
Хлорид магния 0,61
Сульфат натрия 0,071
Фосфат натрия однозамещенный (кристаллический) 0,14
Бикарбонат натрия 0,64
Мочевина 21.0
Глюкоза 0,9
Примечания:
1. Перфузат насыщают той же газовой смесью. 2.рН 7,6.
В таких растворах изолированное сердце рыб сохраняет физиологические свойства и функционирует очень долго. При выполнении простых манипуляций с сердцем допускается использование изотонического раствора хлорида натрия. Однако не стоит при этом рассчитывать на продолжительную работу сердечной мышцы.
§33. КРУГ КРОВООБРАЩЕНИЯ
У рыб, как известно, один круг кровообращения. И, тем не менее, кровь по нему циркулирует дольше. На полный кругооборот крови у рыб уходит около 2 мин (у человека через два круга кровообращения кровь проходит за 20-30 с). Из желудочка через артериальную луковицу или артериальный конус кровь поступает в так называемую брюшную аорту, отходящую от сердца в краниальном направлении к жабрам (рис. 7.3).
Брюшная аорта делится на левые и правые (по количеству жаберных дуг) приносящие жаберные артерии. От них к каждому жаберному лепестку отходит лепестковая артерия, а от нее к каждому лепесточку отходят две артериолы, которые формируют капиллярную сеть из тончайших сосудов, стенка которых образована однослойным эпителием с большими Межклеточными пространствами. Капилляры сливаются в единую выносящую артериолу (по количеству лепесточков). Выносящие артериолы формируют выносящую лепестковую артерию. Лепестковые артерии образуют левую и правую выносящие жаберные артерии, по которым течет артериальная кровь.
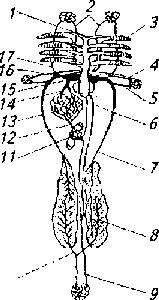
Рис. 7.3. Схема кровообращения костистой рыбы:
1- брюшная аорта; 2 - сонные артерии; 3 - жаберные артерии; 4- подключичные артерия и вена; б- спинная аорта; 7- задняя кардинальная вена; 8- сосуды почек; 9- хвостовая вена; 10 - оборотная вена почек; 11 - сосуды кишечника, 12- воротная вена; 13 - сосуды печени; 14- печеночные вены; 15- венозный 16- кювьеров проток; 17- передняя кардинальная вена
От выносящих жаберных артерий к голове отходят сонные артерии. Далее жаберные артерии сливаются с образованием единого крупного сосуда - спинной аорты, которая тянется по всему телу под позвоночником и обеспечивает артериальное системное кровообращение. Основными отходящими артериями являются подключичная, брыжеечная, подвздошная, хвостовая и сегментарные.
Венозную часть круга начинают капилляры мышц и внутренних органов, которые, объединяясь, формируют парные передние и парные задние кардинальные вены. Кардинальные вены, объединяясь с двумя печеночными венами, образуют кювьеровы протоки, впадающие в венозный синус.
Таким образом, сердце рыб нагнетает и насасывает только венозную кровь. Однако все органы и ткани получают артериальную кровь, так как перед заполнением микроциркуляторного русла органов кровь проходит через жаберный аппарат, в котором осуществляется обмен газов между венозной кровью и водной средой.
§34. ДВИЖЕНИЕ КРОВИ И КРОВЯНОЕ ДАВЛЕНИЕ
Кровь движется по сосудам вследствие разницы ее давления в начале круга кровообращения и в его конце. При измерении кровяного давления без анестезии в вентральном положении (вызывает брадикардию) у лосося в брюшной аорте оно составило 82/50 мм рт. ст., а в дорзальной 44/37 мм рт. ст. Исследование анестезированных рыб нескольких видов показало, что анестезия существенно снижает систолическое давление - до 30-70 мм рт. ст. Пульсовое давление при этом по видам рыб колебалось от 10 до 30 мм рт. ст. Гипоксия приводила к повышению пульсового давления до 40 мм рт. ст.
В конце круга кровообращения давление крови на стенки сосудов (в кювьеровых протоках) не превышало 10 мм рт. ст.
Наибольшее сопротивление току крови оказывает жаберная система с ее длинными и сильно разветвленными капиллярами. У карпа и форели разница систолического давления в брюшной и дорзальной аортах, т. е. при входе и на выходе из жаберного аппарата, составляет 40-50 %. При гипоксии жабры оказывают еще большее сопротивление току крови.
Помимо сердца продвижению крови по сосудам способствуют и другие механизмы. Так, дорзальная аорта, имеющая форму прямой трубы со сравнительно жесткими (по сравнению с брюшной аортой) стенками, оказывает незначительное сопротивление току крови. Сегментарная, каудальная и другие артерии имеют систему кармашковых клапанов, аналогичную тем, которые есть у крупных венозных сосудов. Эта система клапанов препятствует обратному току крови. Для венозного тока крови большое значение имеют также сокращения прилегающих к венам мыши, которые проталкивают кровь в кардиальном направлении.
Венозный возврат и сердечный выброс оптимизируются мобилизацией депонированной крови. Экспериментально доказано, что у форели мышечная нагрузка приводит к уменьшению объема селезенки и печени.
Наконец, движению крови способствуют механизм равномерного наполнения сердца и отсутствие резких систолическо-диастолических колебаний сердечного выброса. Наполнение сердца обеспечивается уже при диастоле желудочка, когда создается некоторое разрежение в перикардиальной полости и кровь пассивно заполняет венозный синус и предсердие. Систолический удар демпфируется артериальной луковицей, имеющей эластичную и пористую внутреннюю поверхность.
Глава 8. ГАЗООБМЕН РЫБ
Концентрация кислорода в водоеме - самый неустойчивый многократно меняющийся в течение суток показатель среды обитания рыб. Тем не менее парциальное давление кислорода и углекислого газа в крови рыб достаточно стабильно и относится к жестким константам гомеостаза.
Как дыхательная среда вода уступает воздушной среде (табл. 8.1).
8.1. Сравнение воды и воздуха как среды дыхания (при температуре 20 њС)
| Показатели
| Воздух
| Вода
| Вода/воздух
| | Плотность, г/см3
| 0,00129
| 1,0
|
| | Вязкость, Па*с
| 0,000018
| 0,0011
|
| | Коэффициент диффузии О2, см2/с
| 0,22
| 0,000021
|
| | Содержание О2, мг/л
|
| 9,2
| 1/30
|
При столь невыгодных изначальных условиях для газообмена эволюция пошла по пути создания дополнительных механизмов газообмена у водных животных, которые позволяют им переносить опасные колебания концентрации кислорода в окружающей их среде. Помимо жабр у рыб в газообмене принимают участие кожа, желудочно-кишечный тракт, плавательный пузырь, специальные органы
§35. ЖАБРЫ - ЭФФЕКТИВНЫЙ ОРГАН ГАЗООБМЕНА В ВОДНОЙ СРЕДЕ
Основная нагрузка в обеспечении организма рыб кислородом и удалении из него углекислого газа ложится на жабры. Они выполняют тетаническую работу. Если сравнивать жаберное и легочное дыхание, то приходишь к заключению, что рыбе необходимо прокачивать через жабры дыхательной среды в 30 раз больше по объему и в 20 000 (!) раз больше по массе.
Более пристальное изучение показывает, что жабры хорошо приспособлены к газообмену в водной среде. Кислород переходит в капиллярное русло жабр по градиенту парциального давления, который у рыб составляет 40-100мм рт. ст. Такова же причина перехода кислорода из крови в межклеточную жидкость в тканях.
Здесь градиент парциального давления кислорода оценивается в 1?15ммрт. ст., градиент концентрации углекислого газа - в 3-15 мм.рт.ст.
Газообмен в других органах, например через кожу, осуществляется по тем же физическим законам, однако интенсивность диффузии в них гораздо ниже. Жаберная поверхность в 10-60 раз превышает площадь тела рыбы. К тому же жабры, высокоспециализированные на газообмене органы, даже при одинаковой с другими органами площади будут иметь большие преимущества.
Самое совершенное строение жаберного аппарата характерно для костистых рыб. Основой жаберного аппарата являются 4 пары жаберных дуг. На жаберных дугах располагаются хорошо васкуляризированные жаберные лепестки, образующие дыхательную поверхность (рис. 8.1).
По стороне жаберной дуги, обращенной в ротовую полость, располагаются более мелкие структуры - жаберные тычинки, которые в большей мере отвечают за механическую очистку воды по мере ее поступления из ротовой полости к жаберным лепесткам.
Поперечно жаберным лепесткам располагаются микроскопические жаберные лепесточки, которые и являются структурными элементами жабр как органов дыхания (см. рис. 8.1; 8.2). Покрывающий лепесточки эпителий имеет клетки трех типов: респираторные, слизистые и опорные. Площадь вторичных ламелей и, следовательно, дыхательного эпителия зависит от биологических особенностей рыбы - образа жизни, интенсивности основного обмена, потребности в кислороде. Так, у тунца при массе 100 г площадь жаберной поверхности составляет 20-30 см2/г, у кефали - 10 см2/г, у форели - 2 см2/г, У плотвы - 1 см2/г.
Жаберный газообмен может быть эффективным только при постоянном токе воды через жаберный аппарат. Вода орошает жаберные лепестки постоянно, и этому способствует ротовой аппарат. Вода устремляется из ротовой полости к жабрам. Такой механизм есть у большинства видов рыб.
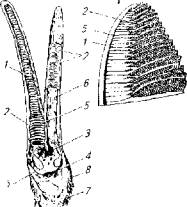
Рис. 8.1. Строение жабр костистых рыб:
1- жаберные лепестки; 2- жаберные лепесточки; 3-жаберная артерия; 4 -жаберная вена; 5-лепестковая артерия; 6 - лепестковая вена; 7-жаберные тычинки; 8- жаберная дуга
Однако известно, что крупные и активные виды, например тунец, рта не закрывают, и дыхательные движения жаберных крышек у них отсутствуют. Такой тип вентиляции жабр называют "таранным"; он возможен только при больших скоростях перемещения в воде.
Для прохождения воды через жабры и движения крови по сосудам жаберного аппарата характерен противоточный механизм, обеспечивающий очень высокую эффективность газообмена. Пройдя через жабры, вода теряет до 90 % растворенного в ней кислорода (табл. 8.2).
8.2. Эффективность извлечения кислорода из воды разными вилами рыб, %
| Вид рыб
| Эффективность дыхания, %
| | Карп
|
| | Камбала
|
| | Форель
|
| | Тунец
|
|
Жаберные лепестки и лепесточки расположены очень тесно, но благодаря малой скорости движения через них воды они не создают большого сопротивления току воды. Согласно расчетам, несмотря на большой объем работы по перемещению воды через жаберный аппарат (не менее 1 м3 воды на 1 кг живой массы в сутки), энергетические затраты рыбы при этом невелики.
Нагнетание воды обеспечивают два насоса - ротовой и жаберный. У разных видов рыб возможно превалирование одного из них. Например, у быстроходных кефали и ставриды действует в основном ротовой насос, а у медлительных придонных рыб (камбалы или сома) - жаберный насос.
Частота дыхательных движений у рыб зависит от многих факторов, но наибольшее влияние на этот физиологический показатель оказывают два - температура воды и содержание в ней кислорода. Зависимость частоты дыхания от температуры показана на рис. 8.2.
Таким образом, жаберное дыхание следует рассматривать как очень эффективный механизм газообмена в водной среде с точки зрения эффективности извлечения кислорода, а также энергозатрат на этот процесс. В том случае, когда жаберный механизм не справляется с задачей адекватного газообмена, включаются другие (вспомогательные) механизмы.
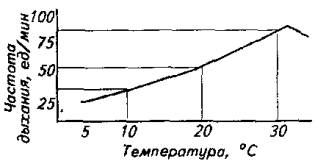
Рис. 8.2. Зависимость частоты дыхания от температуры воды у сеголетков карпа
§36. КОЖНОЕ ДЫХАНИЕ
Кожное дыхание развито в разной мере у всех животных, но у некоторых видов рыб оно может быть основным механизмом газообмена.
Кожное дыхание имеет существенное значение для видов, ведущих малоподвижный образ жизни в условиях низкого содержания кислорода или на короткое время покидающих водоем (угорь, илистый прыгун, сомы). У взрослого угря кожное дыхание становится основным и достигает 60% общего объема газообмена
8.3. Доля кожного дыхания у разных видов рыб
| Вид рыб
| Масса, г
| Температура, њС
| Кожное дыхание, %
| | Угорь
| 100-600
| 13-16
| 21 (до 60)
| | Сомик
|
|
|
| | Карп
| 20-400
| 8-11
| 11-24
| | Карась
|
|
|
| | Осетр
| 90-210
| 18-22
|
| | Плотва
| 40-240
|
|
| | Налим
| 100-300
| 10-12
|
| | Окунь
| 70-370
| 14-18
|
| | Сиг
| 175-200
| 12-13
|
|
Изучение онтогенетического развития рыб свидетельствует о том, что кожное дыхание первично по отношению к жаберному. Эмбрионы и личинки рыб осуществляют газообмен с окружающей средой через покровные ткани. Интенсивность кожного дыхания усиливается с повышением температуры воды, так как повышение температуры усиливает обмен веществ и снижает растворимость кислорода в воде.
В целом интенсивность кожного газообмена определяется морфологией кожи. У угря кожа имеет гипертрофированные по сравнению с другими видами васкуляризацию и иннервацию.
У других видов, например у акул, доля кожного дыхания незначительна, но и кожа у них имеет грубое строение со слаборазвитой системой кровоснабжения.
Площадь кровеносных сосудов кожи у разных видов костистых рыб составляет от 0,5 до 1,5 см:/г живой массы. Соотношение площади кожных капилляров и капилляров жабр варьирует в широких пределах - от 3:1 у вьюна до 10:1 у карпа.
Толщина эпидермиса, колеблющаяся от 31-38 мкм у камбалы До 263 мкм у угря и 338 мкм у вьюна, определяется количеством и размером мукозных клеток. Однако есть рыбы с очень интенсивным газообменом на фоне ординарной макро- и микроструктуры кожи.
В заключение надо подчеркнуть, что механизм кожного дыхания у животных изучен явно недостаточно. Важную роль в этом процессе играет кожная слизь, в составе которой обнаруживается и гемоглобин, и фермент карбоангидраза.
§37. КИШЕЧНОЕ ДЫХАНИЕ
В экстремальных условиях (гипоксия) кишечное дыхание используется многими видами рыб. Однако есть рыбы, у которых желудочно-кишечный тракт претерпел морфологические изменения с целью эффективного газообмена. При этом, как правило длина кишки увеличивается. У таких рыб (сомик, пескарь) воздух заглатывается и перистальтическими движениями кишечника направляется в специализированный отдел. В этой части желудочно-кишечного тракта стенка кишки приспособлена к газообмену, во-первых, за счет гипертрофированной капиллярной васкуляризации и, во-вторых, за счет наличия респираторного цилиндрического эпителия. Заглоченный пузырек атмосферного воздуха в кишке находится под определенным давлением, что повышает коэффициент диффузии кислорода в кровь. В этом месте кишка обеспечивается венозной кровью, поэтому возникают хорошая разница парциального давления кислорода и углекислого газа и однонаправленность их диффузии. Кишечное дыхание широко распространено у американских сомиков. Среди них есть виды с приспособленным для газообмена желудком.
Плавательный пузырь не только обеспечивает рыбе нейтральную плавучесть, но и играет определенную роль в газообмене. Он бывает открытым (лососевые) и закрытым (карп). Открытый пузырь связан воздушным протоком с пищеводом, и его газовый состав может быстро обновляться. В закрытом пузыре изменение газового состава происходит только через кровь.
.В стенке плавательного пузыря имеется особая капиллярная система, которую принято называть "газовой железой". Капилляры железы образуют круто изогнутые противоточные петли. Эндотелий газовой железы способен выделять молочную кислоту и тем самым локально изменять рН крови. Это, в свою очередь, заставляет гемоглобин отдавать кислород прямо в плазму крови. Получается, что кровь, оттекающая от плавательного пузыря, перенасыщена кислородом. Однако противоточный механизм кровотока в газовой железе приводит к тому, что этот кислород плазмы диффундирует в полость пузыря. Таким образом, пузырь создает запас кислорода, который используется организмом рыбы в неблагоприятных условиях.
Другие приспособления для газообмена представлены лабиринтом (гурами, лялиус, петушок), наджаберным органом (рисовый угорь), легкими (двоякодышащие), ротовым аппаратом (окунь ползун), глоточными полостями (Ophiocephalus sp.). Принцип газообмена в этих органах такой же, как в кишке или в плавательном пузыре. Морфологическая основа газообмена в них -это видоизмененная система капиллярного кровообращения плюс утончение слизистых оболочек (рис. 8.3).
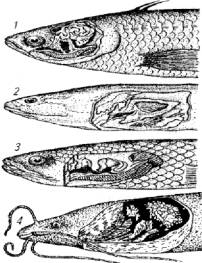
Рис. 8.3. Разновидности наджаберных органов:
1- окунь-ползун: 2- кучия; 3- змееголов; 4- нильский шармут
Морфологически и функционально с органами дыхания связаны псевдобранхии - особые образования жаберного аппарата. Их роль до конца не изучена. То. что к этим структурам притекает кровь от жабр, насыщенная кислородом, свидетельствует о том. что они не участвуют в обмене кислорода. Однако наличие большого количества карбоангидразы на мембранах псевдобранхии допускает участие этих структур в регуляции обмена углекислого газа в пределах жаберного аппарата.
Функционально с псевдобранхиями связана так называемая сосудистая железа, расположенная на задней стенке глазного яблока и окружающая зрительный нерв. Сосудистая железа имеет сеть капилляров, напоминающую таковую в газовой железе плавательного пузыря. Есть точка зрения, что сосудистая железа обеспечивает снабжение сильно насыщенной кислородом кровью сетчатки глаза при максимально низком поступлении в нее углекислого газа. Вероятно, что фоторецепция требовательна к рН растворов, в которых она осуществляется. Поэтому систему псевдобранхии - сосудистая железа можно рассматривать как дополнительный буферный фильтр сетчатки глаза. Если принять во внимание, что наличие этой системы не связано с таксономическим положением рыб, а скорее связано со средой обитания (эти органы имеются чаще у морских видов, обитающих в воде с высокой прозрачностью, и зрение у которых является важнейшим каналом связи с внешней средой), то данное предположение выглядит убедительным.
§38. ПЕРЕНОС ГАЗОВ КРОВЬЮ
Принципиальных отличий в транспортировании газов кровью У рыб нет. Как у легочных животных, у рыб транспортные функции крови реализуются за счет высокого сродства гемоглобина к кислороду, сравнительно высокой растворимости газов в плазме крови, химической трансформации углекислого газа в карбонаты и бикарбонаты.
Основным транспортировщиком кислорода в крови у рыб выступает гемоглобин. Небезынтересно, что гемоглобин рыб функционально делится на два типа- чувствительный к кислоте и нечувствительный к кислоте.
Чувствительный к кислоте гемоглобин при понижении рН крови утрачивает способность связывать кислород.
Нечувствительный к кислоте гемоглобин не реагирует на величину рН, причем для рыб его наличие имеет жизненно важное значение, так как их мышечная активность сопровождается большими выбросами в кровь молочной кислоты (естественный результат гликолиза в условиях постоянной гипоксии).
У некоторых арктических и антарктических видов рыб гемоглобина в крови нет вообще. В литературе есть сообщения о таком же явлении у карпа. В экспериментах на форели показано, что рыба не испытывает асфиксии без функционального гемоглобина (весь гемоглобин искусственно связывали при помощи СО) при температуре воды ниже 5 њС. Это свидетельствует о том, что потребность рыб в кислороде значительно ниже, чем у наземных животных (особенно при пониженных температурах воды, когда повышается растворимость газов в плазме крови).
В определенных условиях с транспортированием газов справляется одна плазма. Однако в обычных условиях у подавляющего большинства рыб газообмен без гемоглобина практически исключен. Диффузия кислорода из воды в кровь протекает по градиенту концентрации. Градиент сохраняется, когда растворенный в плазме кислород связывается гемоглобином, т.е. диффузия кислорода из воды идет до полного насыщения гемоглобина кислородом. Кислородная емкость крови колеблется от 65 мг/л у ската до 180 мг/л у лосося. Однако насыщение крови углекислотой (диоксидом углерода) может снизить кислородную емкость крови рыб в 2 раза.
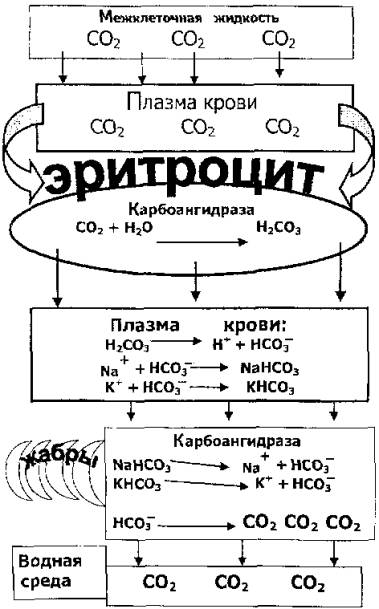
Рис. 8.4. Роль карбоангидразы в переносе углекислого газа кровью
Транспортирование углекислого газа кровью осуществляется по-другому. Роль гемоглобина в переносе углекислого газа в виде карбогемоглобина невелика. Расчеты показывают, что гемоглобин переносит не более 15 % углекислого газа, образующегося в результате обмена веществ рыбы. Основной транспортной системой для переноса углекислого газа является плазма крови.
Попадая в кровь в результате диффузии из клеток, углекислый газ вследствие его ограниченной растворимости создает повышенное парциальное давление в плазме и таким образом должен тормозить переход газа из клеток в кровяное русло. На самом деде этого не происходит. В плазме под влиянием карбоангидразы эритроцитов осуществляется реакция
СО2 + Н2О>Н2СО3>Н+ + НСО3
За счет этого парциальное давление углекислого газа у клеточной мембраны со стороны плазмы крови постоянно снижается, и диффузия углекислого газа в кровь протекает равномерно. Схематично роль карбоангидразы показана на рис. 8.4.
Образующийся бикарбонат с кровью поступает в жаберный эпителий, который также содержит карбоангидразу. Поэтому в жабрах происходит преобразование бикарбонатов в углекислый газ и воду. Далее по градиенту концентрации СО2 из крови диффундирует в омывающую жабры воду.
Протекающая через жаберные лепестки вода контактирует с жаберным эпителием не более 1 с, поэтому градиент концентрации углекислого газа не изменяется и он с постоянной скоростью покидает кровеносное русло. Примерно по такой же схеме происходит удаление углекислого газа и в других органах дыхания. Кроме того, значительные количества углекислого газа, образующегося в результате обмена веществ, выделяются из организма в виде карбонатов с мочой, в составе панкреатического сока, желчи и через кожу.
Дата добавления: 2014-12-11 | Просмотры: 1667 | Нарушение авторских прав
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 34 | 35 |
|