|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
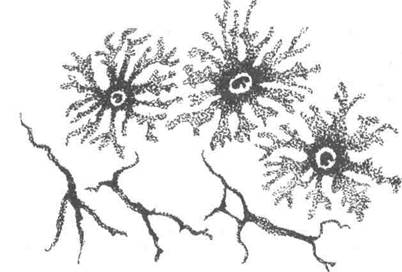
Спеціальні органелиВійки та джгутики - є спеціальними органелами руху Війки роз-ташовані на апікальному полюсі епітеліальних клітин дихальних шля-хів і маткових труб, а джгутики входять до складу сперматозоїдів. Довжина війок досягає 5-10 мкм, джгутиків до 150 мкм. Вони Розділ 1 Основи загальної цитології утворені виростами цитоплазми всередині яких є осьова нитка - аксонема. Аксонема має циліндричну форму і утворена дев'ятьма дуплетами мікротрубочок і двома центральними. Базальне тільце по будові поді-бно до центріолі і структурно зв'язане з аксонемою в єдину систему. Основні білки війок це тубулін та динеїн. Останньому притаманна АТФ-азна активність. Війки рухаються різноманітно: маятникоподіб-но, хвильоподібно, лійкоподібно тощо. Будова джгутиків описана в курсі ембріології. Мікроворсинки це вирости цитоплазми, всередині яких знаходяться скоротливі мікрофіламенти. Останні збільшують апікальну поверхню клітини. Ця органела добре розвинена в епітеліоцитах слизової обо-лонки кишечника. Тонофібрили, міофібрили і нейрофібрили. Ці різновиди мікрофібрил притаманні клітинам та симпластам окремих тканин. Зокрема тонофі-брили та нейрофібрили відповідно епітеліальним та нервовим кліти-нам. Міофібрили забезпечують скорочення скелетних м'язових воло-кон і кардіоміоцитів. Включення Включення це необов'язкові компоненти цитоплазми, котрі утво-рюються і зникають в залежності від метаболічного стану клітини. Розрізняють: трофічні, секреторні, пігментні, вітамінні та екскретор-ні включення. До трофічних включень належать жири, білки і вуглеводи. Напри-клад, нейтральний жир в ліпоцитах пухкої сполучної тканини, гліко-ген в м'язовій тканині та клітинах печінки. Секреторні включення - це округлі утворення різних розмірів, ко-трі містять біологічно-активні речовини. Пігментні включення можуть бути екзогенного походження (ка-ротин, барвники, часточки пилу) і ендогенного (гемоглобін, гемоси-дерин, білірубін, меланін, ліпофусцин, тощо). Наприклад меланоцити шкіри захищають організм від дії ультрафіолетового випромінювання. Присутність пігментів може змінювати колір клітин і тканин. Не рідко пігментація може бути діагностичною ознакою. (рис. 12). Екскреторні включення є продуктами метаболізму які шкідливі для клітин.
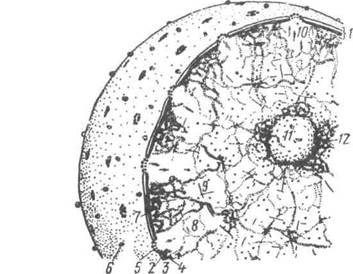
Рис. 12. Шгментні включення у меланоцитах (тотальний нефарбований препарат х 400) Ядро Ядро (від лат. писіеиз і гр. сагіоп) є однією з важливих складових частин клітини. Воно забезпечує збереження і підтримку спадкової ін-формації у вигляді незмінної структури ДНК. В ядрі відбувається від-творення або редуплікація молекул ДНК, що дає змогу при мітозі двом дочірнім клітинам одержувати цілком однакові в якісних і кількісних відношеннях об'єми генетичної інформації. Другою функцією ядра є утворення власне апарата білкового син-тезу. Це не лише синтез, транскрипція на молекулах ДНК різних ін-формаційних РНК, а й транскрипція усіх видів транспортних і рибо-сомних РНК. У ядрі також утворюються субодиниці рибосом шляхом комплексування синтезованих в ядерці рибосомних РНК з рибосом-ними білками, які синтезуються в цитоплазмі і переносяться в ядро (рис. 13). Розділ 1 Основи загальног цитології Ядро може перебувати у мітотичному стані — під час поділу кліти-ни та в інтерфазному — між поділами — метаболічні ядра. У живій клі-тині інтерфазне ядро оптично пусте, в ньому видно лише ядерце. При дії різних пошкоджуючих агентів клітина набуває стану паранекрозу. 3 цього стану клітина може повернутися до нормальної життєдіяль-ності або загинути. В ядрі у цей час морфологічно розрізняють зміни, характерні для загибелі клітини: каріопікноз — ущільнення, каріорек-сис — розпад, каріолізис — розчинення.
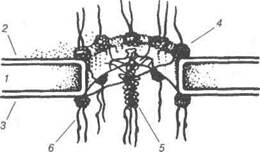
Рис.13. Схема інтерфазного ядра за Заварзіним і Хазаровою: 1-поверхневий апарат ядра; 2-3— мембрани ядерної оболонки (2-зовніш-ня, 3-внутрішня; між ними перинуклеарний простір); 4-щільна пластинка; 5-комлекс з порами; 6-рибосоми; 7-гетерохроматин; 8-еухроматин (ДНП); 9-елементи ядерного матрикса; 10-РНП-частинки; 11-ядерце; 12-навколо-ядерцевий хроматин. Таким чином, ядро являє собою одну з важливих складових частин клітини. Усі клітини тварин містять ядро, за винятком зрілих клітин крові — еритроцитів. Більшість клітин має одне ядро, винятком є дво-ядерні клітини (25-30%) печінки великої рогатої худоби, кролів, білих щурів і свиней. У покривному шарі перехідного епітелію зустрічаються В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія триядерні клітини, а багатоядерні — в остеокластах кісткової тканини, а також в нервових клітинах гангліїв иростати. Форма ядра різна, однак в більшості випадків вона відповідає формі клітини. Так, ядро лімфоцита кулясте, клітин гладеньких м'язів — паличкоподібне, гепатоцитів — ку-лясте, ядро жирових клітин під впливом жирових накопичень сплющене та відтиснене до плазмолеми. У слинних залозах, клітини яких продуку-ють слизовий секрет, ядро також сплющене і розміщується у базальній частині клітини. Ядро лейкоцитів, крім кулястого, може бути сегменто-ваним, бобоподібним, паличкоподібним, мати вигляд підкови. Ядро та цитоплазма — єдина інтегрована система, що знаходиться у постійній рівновазі. Об'єм ядра і цитоплазми кожного типу клітин має своє постійне співвідношення. До складу ядра входять: ядерна оболонка, хроматин, ядерце та ка-ріоплазма. Ядерна оболонка — нуклеолема складається із зовнішньої і вну-трішньої біологічних мембран, відокремлених перинуклеарним про-стором завширшки 20-60 нм. Кожна з мембран має товщину до 8 нм і морфологічно подібна до інших клітинних мембран. Ядерна мембрана відокремлює вміст ядра від цитоплазми, зовнішня — безпосередньо контактує з цитоплазмою клітини. Вона має ряд структурних особли-востей, що дає змогу віднести її до мембранної системи ендоплазма-тичної сітки. На зовнішній мембрані з боку гіалоплазми знаходяться полірибосоми, а сама зовнішня ядерна мембрана переходить в мемб-рани ендоплазматичної сітки. Внутрішня мембрана пов'язана з хромо-сомним матеріалом ядра. Ядерна мембрана містить ядерні пори, утворені в результаті злиття зовнішньої та внутрішньої ядерних мембран. При цьому утворюються округлі перфорації до 90 нм. Вони заповнені складноорганізованими глобулярними та фібрилярними структурами, які разом з мембранною перфорацією утворюють комплекс пори. Він побудований з трьох ря-дів гранул по вісім штук у кожному ряді, діаметр гранул 25 нм. Від гранул відходять фібрилярні відростки. Фібрили, що відходять від пе-риферійних гранул, можуть сходитися в центрі і утворювати своєрідні перегородки, так звані діафрагми пор. Розмір ядерних пор у кожно-го виду клітин є величиною сталою. Кількість ядерних пор залежить від метаболічної активності клітин, густина їх на поверхні нуклеолем більше у клітин з високою метаболічною активністю (рис. 14). Розділ 1 Основи загальної цитології
Рис. 14. Тонка організація ядерної пори: 1-перинуклеарний простір; 2-внутрішня ядерна мембрана; 3-зовнішня ядер-на мембрана; 4-периферійні субодиниці; 5-центральна гранула; 6-фібрили, що відходять від гранул. Ядерна оболонка виконує бар'єрну функцію, відокремлює вміст ядра, його генетичний матеріал від цитоплазми, обмежує вільний до-ступ в ядро та вихід з нього різних речовин, регулює транспорт макро-молекул між ядром і цитоплазмою. Ядерна оболонка бере участь у створенні внутрішньоядерного порядку шляхом фіксації хромосомно-го матеріалу в інтерфазі до внутрішньої ядерної мембрани. Хроматин (від лат. скготаіипит — забарвлення). Завдяки здат-ності сприймати фарбники, ця складова частина інтерфазного ядра названа «хроматин» (Флемінг, 1880). Здатність хроматину сприйма-ти лужні фарбники свідчить про його кислотні властивості, які ви-значаються тим, що до складу хроматину входить ДНК у комплексі з білками. Властивостями фарбуватися володіють і хромосоми, що спостерігається під час мітотичного ділення клітин. Таким чином, хроматин інтерфазних ядер являє собою хромосоми, які у цей час втрачають свою компактну форму, розпушуються, деконденсують-ся. Зону повної деконденсації і її ділянки мають назву еухроматин. При неповному розпушуванні хромосом в інтерфазному ядрі видно ділянки конденсованого хроматину, який називають гетерохромати-ном. Під час мітозу весь еухроматин конденсується і входить до його складу хромосом. До складу хроматину входять складні комплекси дезоксирибону-клеопротеїдів, що складаються з ДНК і спеціальних хромосомних біл- В. П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія ків — гістонів. Хроматин також містить РНК. В кількісному співвідно-шенні ДНК, білок та РНК становлять 1:1,3:0,2. Гістони — лужні білки, збагачені лізином і аргініном. Вони забез-печують специфічну укладку хромосомної ДНК і беруть участь у регу-ляції транскрипції. Гістони розміщені вздовж молекул ДНК не рівномірно, а у вигляді блоків. У один такий блок входить вісім молекул гістонів, які утворюють нуклеосоми, розмір яких 10 нм. При утворенні нуклеосом відбувається компактизація, надспіралізація ДНК, що призводить до скорочення до-вжини хромосомної фібрили у п'ять разів. Саме хромосомна фібрила має вигляд нитки намиста. Такі фібрили додатково повздовжньо кон-денсуються і утворюють основну елементарну фібрилу хроматину тов-щиною 25 нм. Негістонові білки інтерфазних ядер утворюють всереди-ні ядра сітку, яку називають ядерною білковою матрицею. Вона являе собою основу, що визначає морфологію і метаболізм ядра. Хроматин є структурним аналогом хромосом, які можна спостері-гати під час поділу клітин. В деяких випадках ціла хромосома в період інтерфази може залишатися у конденсованому стані, вона має вигляд грудочки гетерохроматину. Наприклад, одна з Х-хромосом у соматич-них клітинах самки залишається у конденсованому стані на стадії дро-блення зиготи. Вперше цей хроматин був досліджений у ядрі клітини кішки і описаний М.Барром і Л.Бертрамом у 1949 р. Його назвали ста-тевим хроматином, або тільцем Барра. Визначення статевого хромати-ну використовують для встановлення генетичної статі організму. В ядрах, крім хроматинових ділянок і матриксу, мають місце пе-рихроматинові фібрили, перихроматинові та інтерхроматинові грану-ли, які містять РНК. Матрицями для синтезу РНК є різні гени, розмі-щені по деконденсованих ділянках хроматинових фібрил. У хроматині міститься 1% ліпідів, роль їх не визначено. Ядерце є найщільнішою структурою ядра (в 1.5 рази перевищує щільність ядра) і добре забарвлюється основними барвниками. Це пов'язано з наявністю в ньому великої кількості РНК, концен-трація якої тут в 2-8 разів, вища ніж у ядрі. Кількість ядерець, як правило, відповідає кількості хромосомних наборів. Тому в диплоїдних клітинах в одному ядрі буває два ядерця. Ядерце - це не самостійна структура, а похідне хромосом, які міс-тять так звані ядерцеві організатори. Останні являють собою локуси Розділ 1 Основи загальної цитології хромосом з найбільш високою концентрацією і активністю синтезу РНК в інтерфазі. Ядерце це місце утворення рибосомних РНК і самих рибосом. Субмікроскопічна будова ядерця представлена в двох основних структурах: гранулах діаметром 15-20 нм і фібрилах товщиною 6-8 нм. Гранули є субодиницями дозріваючих рибосом. Фібрили - це рибону-клеїнові тяжі. Навколо ядерця знаходиться компактна зона навколо-ядерцевого гетерохроматина. Каріоплазма — це рідка частина ядра, в якій містяться ядерні структури. Досліди показали, що після видалення з ядер хроматину і мембран, вони зберігають свою цілісність. Під електронним мікроско-пом встановлено, що до складу каріоплазми входять комплекси пор з фібрилярним периферійним шаром, ядерцеві та численні фібрили, що знаходяться у міжхроматинових районах. Весь комплекс цих структур називають білковим ядерним матриксом. Компоненти матриксу явля-ють собою динамічну структуру; ядерний матрикс відіграє важливу роль в підтримці загальної структури ядра, може брати участь в регу-ляції синтезу нуклеїнових кислот. Дата добавления: 2015-12-15 | Просмотры: 838 | Нарушение авторских прав |