|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Ранні етапи ембріогенезу. Дроблення. ГаструляціяДроблення. В результаті запліднення яйцевої клітини об'єднуються хромосомні набори, настає інтенсивний метаболізм, що характеризуєть- Розділ 2 Ембріологія
у складі зародка з'являється 32 бластомери. На шостому — чотири ме-ридіальних (64 бластомери) і на сьомому — чотири широтні (128 блас-томерів). Утворені бластомери зразу заокруглюються і стикаються з сусідніми бластомерами лише невеликою частиною своєї поверхні.
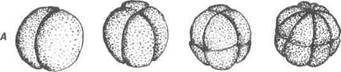
Рис. 26. Повне рівномірне дроблення зиготи: А — дроблення зиготи ланцетника: I — стадія двох бластомерів; II — ста-дія чотирьох бластомерів; III — стадія восьми бластомерів; IV — стадія 16-ти бластомерів; V — стадія 32-х бластомерів; VI — стадія 64-х бластоме-рів; VII — стадія 128 бластомерів; Б — бластула ланцетника: 1 — бластодерма; 2 — бластоцель; 3 — дно бластули; 4 — крайова зона; 5 — дах бластули Бластомери розміщуються не купкою, а в один шар, утворюючи стінку порожнистої кулі. Таким чином, при повному рівномірному дробленні всі бластомери діляться одночасно (синхронно); їх кіль-кість зростає за принципом геометричної прогресіїї. Наступні ділення перестають бути синхронними, темпи ділення більших клітин веге- В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія ся посиленням дихання та синтетичних процесів, зокрема синтезу ДНК, РНК, білків, підвищується активність рибосом, зростає фосфорний, ка-лієвий та кальцієвий обмін, змінюється активність протеолітичних фер-ментів, підвищується проникність мембран, спостерігаються інші зміни. Всі ці метаболічні зрушення стимулюють мітотичне ділення зиготи, за яким настає поділ ядер і цитоплазми. Клітини, що поділилися, не рос-туть, розмір їх зберігається. Кількість ДНК в ядрі подвоюється, після чергового поділу клітини зберігають диплоїдність. Через відсутність росту клітин після поділу, зигота роздрібнюється на все менші клітини. Останні називаються бластомерами (від гр. Біазіоз - зародок, тегоз — частина), а площини, що їх розділяють — борознами дроблення. Таким чином, дроблення — це багаторазові мітотичні поділи зиго-ти, в результаті яких зародок стає багатоклітинним, не змінюючи сут-тєво при цьому свого розміру. Типи дроблення зиготи тісно пов'язані з вмістом у яйцеклітині по-живних речовин. Розрізняють дроблення повне рівномірне, повне нерівномірне та часткове — дискоїдальне. Повне рівномірне дроблення. Класичним прикладом повного рівно-мірного дроблення оліголецитальних яйцеклітин є розвиток ланцет-ника, вперше вивчений О.О.Ковалевським. В заплідненій яйцеклітині розрізняють два полюси: верхній — анімальний і нижній — вегетатив-ний (рис. 26). Після запліднення жовток, незначна кількість якого була рівно-мірно розподілена по всій цитоплазмі, переміщується до вегетативного полюсу. Перша борозна дроблення проходить у меридіональному на-прямку і ділить зиготу на два бластомери, які відповідають майбутній лівій та правій половині тіла зародка. Друга борозна дроблення також проходить меридіонально під прямим кутом до першої. Тепер зародок складається з чотирьох бластомерів. Третя борозна дроблення має ек-ваторіальний напрям, тому кожний бластомер ділиться на дві частини. Такий зародок побудований із восьми бластомерів, при цьому чотири із них утворилися із вегетативного полюсу зиготи, в зв'язку з цим вони містять увесь жовток зиготи і відрізняються великими розмірами. Ці бластомери відповідають задній частині тіла; анімальні (чотири) — пе-редній частині. Потім з'являються дві меридіальні борозни, які ділять зародок на 16 бластомерів, на п'ятому діленні — дві широтні борозни — Розділ 2 Ембріологія Дроблення починається з утворення двох меридіональних борозен дроблення, які настають одна за другою під прямим кутом. Вони скоро ділять позбавлений жовтка анімальний полюс зиготи на два, а потім на чотири бластомери. Вегетативний полюс, що містить увесь жовток зиготи, дробиться значно повільніше, і бластомери, що виникають тут, мають більші розміри. Третя борозна розміщується ближче до анімаль-ного полюса зиготи і має широтний напрямок. Широтні борозни дроблення замінюються меридіональними і при цьому скоро виникає асинхронність та тангенціальність (поділ блас-томєрів у площині, паралельній поверхні зиготи) в дробленні, тому воно закінчується утворенням багатошарової бластули. Дах бласту-ли складається з дрібних бластомерів, які називають мікромерами, а дно — із великих бластомерів — макромерів. Увесь жовток знаходиться в макромерах. Бластоцель зрушений до анімального полюса та змен-шений у розмірах. Бластула, що утворилася в результаті повного (го-лобластичного) дроблення, називається целобластулою. Часткове, або меробластичне (дископодібне), дроблення спостеріга-ється у риб, рептилій, птахів і характерне для полілецитальних і тело-лецитальних яйцеклітин. В полілецитальних яйцеклітинах дроблення охоплює лише вільну від жовтка цитоплазму, де знаходиться ядро, воно не поширюється на вегетативний полюс, останній залишається нероз-діленим на бластомери. Перші дві меридіональні борозни проходять через анімальний полюс під кутом один до одного. Меридіональні бо-розни чергуються з широтними та тангенціальними. Бластомери, що утворилися в процесі дроблення, розміщуються на жовткові в один шар. Останній називають бластодиском, тому дроблення дискоїдальне. Роз-виток зародка відбувається за рахунок зародкового щитка — централь-ної частини зародка. Остання частина зародкового диска бере участь в утворенні тимчасових (провізорних) органів — зародкових оболонок, які створюють сприятливі умови для розвитку зародка. В результаті дроблення утворюється бластула. її бластоцель має вигляд вузької щілини, зрушеної до анімального полюса. Дах бластули складається з бластомерів. Крайова зона — це клітини, що інтенсивно ді-ляться (бластомери). Дном є нерозділений на бластомери жовток вегета-тивного полюса зиготи. Такий тип бластули називають дискобластулою. Гаструляція та утворення осьових органів. Після дроблення та утворення бластули настає важлива стадія розвитку зародка — га- В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія тативного полюсу сповільнюються. Скоро кількість клітин досягає тисячі. Виникає одношаровий зародок — бластула. Стінку бластули називають бластодермою, а заповнену драглистою масою порожни-ну — первинною порожниною тіла — бластоцелем. В бластодермі ви-діляється дно, яке виникло із більших і багатих на жовток клітин на місці колишнього вегетативного полюсу зиготи. Дно обмежене кра-йовою зоною; усі інші клітини бластодерми, які виникли в анімальній половині яйцеклітини, утворюють дах бластули. Прикладом повного нерівномірного дроблення є розвиток мезоле-цитальних яйцеклітин амфібії (рис. 27).
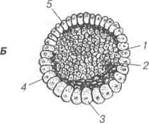
Рис. 27. Схема повного нерівномірного дроблення зиготи амфібії: 1 — мікромери; 2 — макромери; 3 — бластодерма; 4 — бластоцель В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія струляція (§азїег — шлунок). Для гаструляції характерний глибокий та упорядкований розподіл клітин у зародку. Вона характеризується процесами поділу і диференціюванням клітин, їх перегрупуванням, взаємодією та впливом одних на інших. У результаті гаструляції утво-рюється зародок, який спочатку складається з двох, а пізніше з трьох зародкових листків: ектодерми (від гр. екіоз — зовнішній), ентодерми (від гр. епіої — внутрішній) і мезодерми (від гр. тезоз — середній). Із цих зародкових листків шляхом морфогенетичних переміщень утворюються осьові органи: нервова трубка, хордомезодермальний за-чаток (від гр. спогсіа — струна) та кишкова трубка. Поява цих зачат-ків починається на стадії бластули і називається презумптивними (від гр. ргезитрііо — передбачення) зачатками. Так, дах бластули, побудо-ваний із презумптивних зачатків ектодерми та нервової трубки, дно бластули — із презумптивного зачатка — кишкової трубки, а крайова зона — із хордальної пластинки та мезодерми. У вивченні нроцесів гаструляції значна роль належить В.Фогту (1929), який вперше запропонував методику маркування частин зарод-ка. Для вивчення гаструляції були використані різні барвники та радіо-активні ізотопи, завдяки яким досліджено переміщення окремих груп клітин. Більша частина цих досліджень грунтується на використанні прижиттєвих барвників. Виготовлення гістологічних зразків зародків у різні періоди після нанесення мітки на клітини тієї чи іншої локалізації дає змогу простежити, що відбувається з клітинами після того, як вони у процесі гаструляції потрапляють в ті чи інші частини зародка. На ста-дії пізньої бластули амфібії і різних інших тварин можна цілком точно визначати частини, із яких будуть розвиватися ті чи інші органи. Тому прийнято говорити про презумпцію та карти презумптивних органів. Як особливості будови овоцита визначають характер дроблення, так і гаструляція залежить від специфічних для різних класів тварин характеристик їх бластул. Розрізняють чотири різновиди гаструляції: інвагінація (впинання), імміграція (переміщення клітин із зануренням), епіболія (обростання) і делямінація (розщеплення шару на два). У різних хордових гаструля-ція відбувається в поєднанні кількох її різновидів, однак один із них є основним. Так, у ланцетника, яйцеклітина якого оліголецитальна, її дроблен-ня повне рівномірне — гаструляція відбувається шляхом інвагінації. В Розділ 2 Ембріологія
Дорсальна частина ектодерми перетворюється в нервову пластин-ку, а решта ектодерми — епідерміс шкіри. Нервова пластинка шляхом перетворень формує нервову трубку, яка стає джерелом розвитку спинного мозку У хребетних передній кінець нервової трубки утво-рює головний мозок. Поряд з утворенням нервової трубки розвива-ється первинна ентодерма. В її спинній частині під нервовою трубкою з'являється клітинний тяж, який перетворюється у хорду 3 боків від хорди завдяки активному мітозу формується два мішкоподібних ви-рости. Вони є зачатком третього зародкового листка — мезодерми. Остання сегментується і утворює соміти — спинні сегменти, сегментні ніжки та несегментовану частину, яка складається із двох листків: зо-внішнього — паріетального і внутрішнього — вісцерального. Між ними виникає вторинна порожнина тіла — целом. Гаструляція у амфібій (рис. 28, 29) не може відбуватися так, як у ланцетника, оскільки вегетативна півкуля яйця дуже перевантажена жовтком. її дроблення повне нерівномірне й закінчується формуван-ням багатошарової бластули, бластоцель якої зміщений до анімаль-ного полюса. Бластомери анімального нолюса дрібні — мікромери, вегетативного — великі, їх називають макромерами. Тому інвагінація відбувається обмежено, поряд з нею спостерігається епіболія. Гастру-ляція відбувається шляхом інвагінації та епіболії. Активні мікромери напливають на вегетативну половину бластули — макромери, при цьо-му утворюється серпоподібна борозна — дорсальна трубка бластопора (від гр. ьіазіоз - зародок, рогоз — отвір), через неї починає підгинатися прехордальна, потім хордальна пластинки. Складка, що утворюється, 6 - 8-305 В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія являє собою первинну кишку, її порожнина — гастроцель збільшуєть-ся, а бластоцель витісняється і зменшується. В результаті цього тонка клітинна перегородка, що відокремлювала бластоцель від гастроцеля, розривається, обидві порожнини з'єднуються і утворюють порожни-ну первинної кишки. Прехордальна пластинка разом з прехордальною ентодермою прошнуровують дорсальну губу, потім через неї прохо-дить матеріал хорди, що становить дах первинної кишки. Хордальна пластинка перетворюється у хорду і відокремлюється від первинної кишки, остання на дорсальному боці залишається незамкненою. За ра-хунок вільних кінців ентодерми, що ростуть назустріч один другому під зачатком хорди, порожнина первинної кишки замикається. Поряд з матеріалом хорди через бокові губи бластопора, який набуває підко-воподібної форми, в глибину зародку переміщується і матеріал мезо-дерми й розміщується між екто- та ентодермою.
Рис. 28. Гаструляція зародка (А, Б, В, Г — ланцетника; Д, Е, Є — ам фібії): 1 — бластоцель; 2 — гранули жовтка; 3 — ектодерма; 4 — майбутня хорда; 5 — гастроцель; 6 — бластопор; 7 — мезодерма; 8 — ентодерма; 9 — жовток; 10 — бластодерма; 11 — жовткова пробка Розділ 2 Ембріологія 11
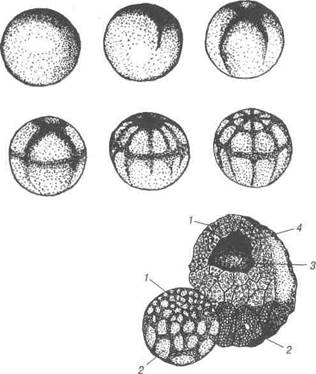
Рис. 29. Стереограма зародка амфібії: 1 — ектодерма; 2 — міотом; 3 — дерматом; 4 — хорда; 5 — паріетальний лис-ток; 6 — вісцеральний листок; 7 — кишка; 8 — сегменти; 9 — сегментна ніжка; 10 — спланхнотом; 11 — мезодерма; 12 — нервова трубка. Вентральна губа бластопора утворюється в результаті вп'ячування вентральної частини мезодерми. Целом у зародка амфібій з'являється в результаті перерозподі-лу клітин мезодермального зачатка на паріетальний та вісцеральний листки. Форма бластопора у амфібій чітко відображає вплив жовтка на процес гаструляції. У амфібій бластопор вузький, він закритий жовт-ковою пробкою — великою кількістю багатих на жовток бластомерів, що становили дно бластули. Майбутній епітелій і матеріал нервової системи покриває всю по-верхню зародка, їх поширення залежить від інтегративного впливу всього зародка. У риб, в зв'язку з перевантаженням їх яйцеклітин жовтком, дро-блення поверхневе, частково меробластичне, в результаті цього утво-рюється дискобластула; її дах утворений зародковим диском, а дно — поверхневим шаром — перибластом, який являє собою жовток, що не дробився. Перибласт містить незначну кількість клітин, що відокреми-лись від бластодиска та ядер надлишку сперміїв. Ферменти цих клітин 6- 83 В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологгя беруть участь у засвоєнні жовтка зародком в перші дні його розвитку. Гаструляція у одних представників хордових відбувається шля-хом міграції — масове переміщенням клітин в напрямку заднього краю бластодиска, внаслідок цього останній стає товстим. Через цей по-товщений край підгинається матеріал. Матеріали майбутньої прехор-дальної ентодерми, прехордальної пластинки і хорди обгинають задній край і рухаються під поверхневим шаром бластодиска в протилежно-му напрямку — вперед, утворюючи первинну кишку. Дах цієї кишки складається з хордальної пластинки, дно — із вп'яченої прехордальної ентодерми та клітин перибласта і частини внутрішніх клітин бласту-ли. В міру того, як цей матеріал входить в середину, на його місці по задньому краю утворюється крайова зарубка. Вона відповідає блас-топору Його середня частина — це дорсальна губа, латерально від неї лежать бокові губи, вентральна губа бластопора відсутня, її роль у риб відіграють бокові краї зародкового диска. Ентодерма відокремлюється від зародкового диска і між остан-ньою та жовтком утворюється порожнина первинної кишки. Хордоме-зодермальний зачаток спочатку входить до складу ентодерми, а потім, після міграції, розміщується між ектодермою та ентодермою. Середня частина мезодермального зачатка — це матеріал майбутньої хорди, з боків її — мезодерма. Клітини зародкового диска не містять жовтка, в результаті цього гаструляція та закладання осьових органів у цих представників хор-дових закінчується дуже швидко. Зародкові листки обростають жов-ток, де знаходяться поживні речовини і формують тимчасовий орган за межами зародка — жовтковий мішок. В його утворенні беруть участь усі три зародкові листки: ектодерма, ентодерма та мезодерма. Внутрішня поверхня жовткового мішка являє собою позазародко-ву ентодерму, вона всмоктує поживні речовини із жовтка, середній шар стінки жовткового мішка становить позазародкова мезодерма, вона утворює густу сітку кровоносних судин, по яких рухаються всмоктані ентодермою поживні речовини в тіло зародка. Позазародкова ектодер-ма являє собою зовнішню оболонку стінки жовткового мішка, яка за-хищає від пошкодження ніжну сітку жовткових кровоносних судин. В подальшому, головний та хвостовий кінці зародка відокремлюють-ся від поверхні жовтка, а навколо його тулуба утворюється тулубова складка, що відокремлює зародок від жовтка, залишаючись пов'язаною Розділ 2 Ембріологія з ним лише жовтковим стебельцем. Після використання поживних ре-човин жовткового мішка останній відпадає, а на місці жовткового сте-бельця залишається лише пупок. 2.6 Розвиток птахів Яйцеві клітини у птахів досить великі і по класифікації полілеци-тальні та телолецитальні. Після закінчення яйцевою клітиною рос-ту, в результаті нагромадження у її цитоплазмі жовтка, відбувається овуляція. Фолікулярний епітелій у тому місці фолікула, що назива-ється стигмою, під впливами м'язового скорочення стінки фолікула, поступово стоншується, потім розривається. Яйцеклітина випадає із фолікула в просвіт лійки яйцепроводу, який охоплює у цей час яєчник. Запліднення відбувається у лійці яйцепроводу. В цей час яйцеклітина знаходиться на стадії другого ділення дозрівання. Для птахів характерна поліспермія, проте з пронуклеусом самки зливається лише один сперматозоїд. Третинні оболонки яйцеклітини утворюються в різних відділах яйцепроводу, кожний із яких спеціа-лізований або на виділення білка, або на утворення підшкаралупних оболонок, чи шкаралупи. Одночасно з формуванням третинних обо-лонок відбувається меробластичне дроблення. У яйцепроводі яйце перебуває 4-27 годин. Залежно від цього різ-ним буває і ступінь розвитку зародка в знесеному яйці. Частіше заро-док знаходиться на стадії бластули або ранньої гаструли. В дробленні бере участь лише анімальний полюс зиготи, де зна-ходиться ядро та цитоплазма, позбавлена жовтка. Вегетативний полюс участі в дробленні не бере, оскільки завантажений масою жовтка, який гальмує дроблення. Перші дві борозни дроблення мають меридіональ-ний напрямок і розміщені перпендикулярно одна до другої. Мериді-альні борозни замінюються широтними (рис. 30). Клітини (бластомери), що утворилися, лежать в жовтку у вигляді диска, тому дроблення називають дискоїдальним, а бластула, що утво-рилася — дискобластула. її дах і крайова зона складається із бластоме-рів диска, а дно — із неподіленого на клітини жовтка. Бластоцель має вигляд вузької щілини, зміщеної до анімального полюса бластули. Цен-тральна частина бластодиска, із якої в подальшому розвивається заро-док, називається зародковим щитком (зародковим диском, бластодис-
ком); по краях він обмежений піднятим над жовтком прозорим валиком, який називають прозорою зоною (від лат. агеа - обідок, реііисісіа - про-зора). Периферійну від неї частину бластодиска, що щільно прилягає до жовтка, називають темною зоною (від лат. агеа - обідок, ораса - темна). Зовнішній край цієї зони є кінцем обростання жовтка бластодиска. Клі-тини кінця обростання знаходяться у тісному зв'язку з жовтком.
А-перше ділення дроблення; Б-друге ділення дроблення; В — третє ділення дроблення; Г — четверте ділення дроблення; Д — п'яте ділення дроблення; Е — рання морула. Розділ 2 Ембріологія Розвиток зародка після знесення яйця затримується, якщо його температура знижується нижче температури тіла птиці. Помірне охо-лодження не обов'язково призводить до загибелі зародка. Дана об-ставина має велике практичне значення в промисловій інкубації яєць сільськогосподарської птиці. Гаструляція. У птахів гаструляція відбувається у дві фази. В пер-шій фазі від бластодиска шляхом делямінації всередину відчленову-ється гіпобласт, який дає початок ентодермі. Поряд з цим верхній шар бластодиска, який називають епібластом, стає зовнішнім зародковим листком — ектодермою. На другій фазі гаструляції від епібласта від-окремлюється матеріал, із якого утворюється хорда та мезодерма. По-тім в складі епібласта залишається лише ектодерма. В подальшому важливі процеси відбуваються в епібласті. По периферії його спостері-гаються складні переміщення клітин, в яких переважають сходження в задній кінець бластодиска, там виникає згущення клітин епібласта у вигляді повздовжнього тяжа, що називається первинною смужкою. Вона розміщена по середній лінії зародкового щитка і спрямовується від заднього краю зародкового щитка вперед. Через деякий час піс-ля виникнення первинна смужка утворює по середній лінії жолобок і первинну ямку на передньому кінці. Первинну смужку з жолобком називають первинною борозенкою, а згущення клітин на передньому кінці первинної бороздки — гензеновським вузликом. Первинна смужка ідентична бластопору гаструли ланцетника та амфібій. Первинна ямка гензеновського вузлика — це гомолог дорсальної губи. Останні зони первинної смужки гомологічні боковим та вентральним губам. У зоні первинної ямки зародковий матеріал впинається у вигляді клітинного тяжа та переміщується між ектодермою та ентодермою до передньо-го краю зародка. Цей тяж називають хордальним відростком, з нього розвивається хорда. Через первинну бороздку в напрямку переднього кінця зародка мігрують два крилоподібні зачатки, що ростуть з боків від хорди між ектодермою та ентодермою. Ці зачатки формують мезо-дерму (рис. 31). У цілому друга фаза гаструляції птахів і інших амніот гомологічна гаструляції амфібій. I у птахів гаструляція закінчується формуванням зародкових шарів. У гаструляції птахів спостерігається подальше удо-сконалення у розвитку хребетних: зачаток хорди та мезодерми повніс-тю втратили зв'язок з первинною кишкою.
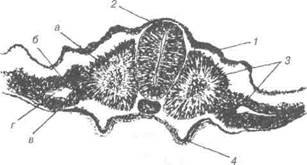
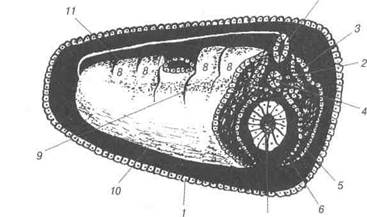
1 — гензенівський вузлик; 2 — світле поле; 3 — темне поле; 4 — первинна ямка; 5 — первинна бороздка; 6 — первинний валик. Подальший розвиток зародкових листків і їх похідних у птахів відбувається як і в інших хордових. Із ектодерми утворюється нерво-ва пластинка, а після з'єднання її країв формується нервова трубка. Остання є матеріалом розвитку всієї нервової системи, а ектодерма — зачаток епідермісу (рис. 32). В мезодермі виділяються соміти — сегментована мезодерма та несегментована мезодерма (спланхнотом). В процесі розвитку кож-ний соміт диференціююєься на дерматом, міотом і склеротом. Із дерматома розвиваються глибокі шари шкіри, із міотома — м'язо-ва тканина скелета, із склеротома — сполучнотканинні структури. Із сегментних ніжок соміта розвивається сечовидільна система. В спланхнотомі виділяються паріетальний (пристінний) і вісцераль-ний (внутрішній) листки, між якими знаходиться целом - первинна порожнина тіла. Розділ 2 Ембріологія
Рис. 32. Зародкові листки та осьові органи курки Дата добавления: 2015-12-15 | Просмотры: 1016 | Нарушение авторских прав |


 бластоцель починають проникати клітини крайовоїзони та великі блас-томери дна бластули. При цьому бластоцель поступово витісняється і із одношарової бластули утворюється двохшарова чаша, яка складається із зовнішнього листка — ектодерми і внутрішнього - ентодерми. Нову порожнину в цій двошаровій чаші називають гастроцелем. Отвір, що веде із гастроцеля назовні, називають бластопором. Його краї утворю-ють чотири губи: дорзальну, вентральну і дві бокових. Через губи блас-топора інвагінує матеріал різних ембріональних зачатків. Так, дорсаль-на губа відмежовує матеріал майбутньої хорди, що увійшов до складу дорсальної частини даху первинної кишки. Бокові та вентральні губи пов'язані з матеріалом майбутньої мезодерми і стінки первинної киш-ки. В кінці гаструляції зародок швидко росте у довжину.
бластоцель починають проникати клітини крайовоїзони та великі блас-томери дна бластули. При цьому бластоцель поступово витісняється і із одношарової бластули утворюється двохшарова чаша, яка складається із зовнішнього листка — ектодерми і внутрішнього - ентодерми. Нову порожнину в цій двошаровій чаші називають гастроцелем. Отвір, що веде із гастроцеля назовні, називають бластопором. Його краї утворю-ють чотири губи: дорзальну, вентральну і дві бокових. Через губи блас-топора інвагінує матеріал різних ембріональних зачатків. Так, дорсаль-на губа відмежовує матеріал майбутньої хорди, що увійшов до складу дорсальної частини даху первинної кишки. Бокові та вентральні губи пов'язані з матеріалом майбутньої мезодерми і стінки первинної киш-ки. В кінці гаструляції зародок швидко росте у довжину.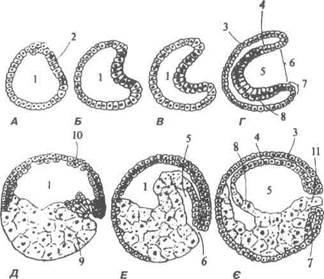

 В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія
В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія
 Рис. 30. Бластодерма яйця голуба на ранніх стадіях дроблення:
Рис. 30. Бластодерма яйця голуба на ранніх стадіях дроблення: В.П. Новак, Ю.П. Бичков, М І< > II" уіііпіко Цитологія, гістологія, ембріологія
В.П. Новак, Ю.П. Бичков, М І< > II" уіііпіко Цитологія, гістологія, ембріологія 'Ч^
'Ч^ Рис. 31. Курячий зародок на стадії первинної смужки після 16 годин інкубації:
Рис. 31. Курячий зародок на стадії первинної смужки після 16 годин інкубації: