|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
ЗАКОНОМЕРНОСТИ РАСПРЕДЕЛЕНИЯ НЕРВОВ1. Соответственно группировке органов тела вокруг нервной системы 2. Соответственно строению тела по принципу двусторонней симметрии 3. Соответственно метамерному строению туловища нервы этой области 4. Нервы идут по кратчайшему расстоянию от места выхода из спин При перемещении органа от места первичной закладки в область окончательного расположения его после рождения нерв равтет и следует за органом. 5. Нервы мышц отходят от сегментов спинного мозга, соответствующих Таким образом, по месту происхождения нерва можно определить область эмбрионального развития органа, ибо существует соответствие между происхождением нерва и местом закладки органов. 6. Если мышца представляет собой продукт слияния нескольких мио- широких мышц живота межреберными нервами и ветвями поясничного сплетения). То же наблюдается в отношении висцеральных мышц, развивающихся из материала нескольких жаберных дуг. Так, переднее брюшко двубрюшной мышцы, возникающее из 1-й жаберной дуги, иннервируется тройничным нервом, а заднее брюшко, производное 2-й жаберной дуги, — лицевым нервом. 7. Поверхностные нервы (кожные) сопровождают подкожные вены, глубо 8. Нервы, заложенные в сосудисто-нервных пучках, как и эти последние, ВЕГЕТАТИВНАЯ (АВТОНОМНАЯ) НЕРВНАЯ СИСТЕМА Выше отмечалась коренная качественная разница в строении, развитии и функции неисчерченных (гладких) и исчерченных (скелетных) мышц. Скелетная мускулатура участвует в реакции организма на внешние воздействия и отвечает на изменение среды быстрыми и целесообразными движениями. Гладкая мускулатура, заложенная во внутренностях и сосудах, работает медленно, но ритмично, обеспечивая течение жизненных процессов организма. Эти функциональные различия связаны с разницей в иннервации: скелетная мускулатура получает двигательные импульсы от анимальной, соматической части нервной системы, гладкая мускулатура — от вегетативной. Вегетативная нервная система управляет деятельностью всех органов, участвующих в осуществлении растительных функций организма (питание, дыхание, выделение, размножение, циркуляция жидкостей), а также осуществляет трофическую иннервацию (И. П. Павлов). Трофическая функция вегетативной нервной системы определяет питание тканей и органов применительно к выполняемой ими функции в тех или иных условиях внешней среды (адаптационно-трофическая функция). Известно, что изменения в состоянии высшей нервной деятельности отражаются на функции внутренних органов и, наоборот, изменение внутренней среды организма оказывает влияние на функциональное состояние центральной нервной системы. Вегетативная нервная система усливает или ослабляет функцию специфически работающих органов. Эта регуляция имеет тонический характер, поэтому вегетативная нервная система изменяет тонус органа. Так как одно и то же нервное волокно способно действовать лишь в одном направлении и не может одновременно повышать и понижать тонус, то сообразно с этим вегетативная нервная система распадается на два отдела, или части: симпатическую и парасимпатическую — pars sympathica и pars para-sympathica. Симпатический отдел по своим основным функциям является трофическим. Он осуществляет усиление окислительных процессов, потребление питательных веществ, усиление дыхания, учащение деятельности сердца, увеличение поступления кислорода к мышцам. Роль парасимпатического отдела охраняющая: сужение зрачка при сильном свете, торможение сердечной деятельности, опорожнение полостных органов. Сравнивая область распространения симпатической и парасимпатической иннервации, можно, во-первых, обнаружить преобладающее значение одного какого-либо вегетативного отдела. Мочевой пузырь, например, получает в основном парасимпатическую иннервацию, и перерезка симпатических нервов не изменяет существенно его функции; только симпатическую иннервацию
Рис. 339. Общая схема вегетативной нервной системы. Пунктиром обозначены постганглионарные волокна симпатической системы, идущие к органам; непрерывной линией — преганглионарные волокна парасимпатической и симпатической систем. Область центров симпатической системы в спинном мозге заштрихована. / — центры парасимпатической системы в головном мозге (краниальный отдел); 2 — центры симпатической системы; 3- центры парасимпатической системы в нижнем конце спинного мозга (крестцовый отдел); 4 — половые органы; 5 — мочевой пузырь; 6 — толстая кишка; 7 - тонкая кишка; 8 - почка; 9 — надпочечник; 10 — поджелудочная железа; 11 - печень; 12 - желудок; 13 -сердце; 14 — легкие; /5 - сосуды головы; 16, 17 — слюнные железы; 18 — глаз; 19 — полосатое тело. получают потовые железы, волоско-вые мышцы кожи, селезенка, надпочечники. Во-вторых, в органах с двойной вегетативной иннервацией наблюдается взаимодействие симпатических и парасимпатических нервов в форме определенного антагонизма. Так, раздражение симпатических нервов вызывает расширение зрачка, сужение сосудов, ускорение сердечных сокращений, торможение перистальтики кишечника; раздражение парасимпатических нервов приводит к сужению зрачка, расширению сосудов, замедлению сердцебиения, усилению перистальтики. Однако так называемый антагонизм симпатической и парасимпатической частей не следует понимать статически, как противопоставление их функций. Эти части взаимодействующие, соотношение между ними динамически меняется на различных фазах функции того или иного органа; они могут действовать и антагонистически, и синергически. Антагонизм и синергизм — две стороны единого процесса. Нормальные функции нашего организма обеспечиваются согласованным действием этих двух отделов вегетативной нервной системы. Эта согласованность и регуляция функций осуществляются корой головного мозга. В этой регуляции участвует и ретикулярная формация. Автономия деятельности вегетативной нервной системы не является абсолютной и проявляется лишь в местных реакциях коротких рефлекторных дуг. Поэтому предложенный PNA термин «автономная нервная система» не- является точным, чем и объясняется сохранение старого, более правильного и логичного термина «вегетативная нервная система». Деление вегетативной нервной системы на симпатический и парасимпатический отделы проводится главным образом на основании физиологических и фармакологических данных, но имеются и морфологические отличия, обусловленные строением и развитием этих отделов нервной системы. Поэтому охарактеризуем сначала морфологические особенности вегетативной нервной системы в сравнении с анимальной. Опишем прежде всего центры вегетативной нервной системы (рис. 339).
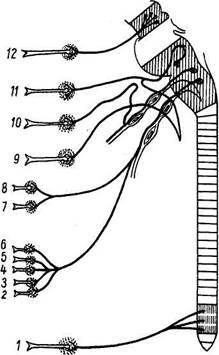
Рис. 340. Схема парасимпатической нервной системы. Преганглионарные волокна изображены сплошными линиями, постганглионарные — стрелками. 1 - подчревное сплетение: парасимпатические преганглионарные волокна подходят в составе тазовых нервов, постганглионарные направляются к прямой кишке, мочевому пузырю, половым органам. Постганглионарные волокна: 2-к почкам; 3-к поджелудочной железе; 4-к печени; 5-к кишечнику; 6-к желудку; 7-к бронхам; 8 — к сердцу (преганглионарные волокна 2—8 подходят в составе блуждающего нерва); 9 — поднижнечелюстной узел: преганглионарные волокна подходят по нерву, носящему название барабанной струны (ветвь лицевого нерва), постганглионарные направляются к поднижнечелюстной и подъязычной слюнным железам; 10 — ушной узел: преганлионарные волокна подходят по ветви языкоглоточного нерва; постганглионарные направляются к околоушной железе, 11 — крылонебный узел: преганглионарные волокна подходят по большому каменистому нерву (ветвь лицевого нерва), постганглионарные направляются к слезной железе и железам полости рта и носа; 12 — ресничный узел: преганглионарные волокна подходят по глазодвигательному нерву, постганглионарные направляются к сфинктеру зрачка и ресничной мышце глаза. Анимальные нервы выходят из мозгового ствола и спинного мозга на всем их протяжении сегментарно, причем эта сегментарность сохраняется частично и на периферии. Вегетативные нервы выходят только из нескольких отделов (очагов) центральной нервной системы. Имеются 4 таких очага, откуда выходят вегетативные нервы: 1. Мезэнцефалический отдел в среднем мозге (nucl. accessorius и непарное 2. Бульварный отдел в продолговатом мозге и мосте (ядра VII, IX и X пар 3. Тораколюмбалъный отдел в боковых рогах спинного мозга на протяже 4. Сакральный отдел в боковых рогах спинного мозга на протяжении Тораколюмбальный отдел относится к симпатической системе, а краниальный и сакральный — к парасимпатической (см. рис. 339; рис. 340). Над этими очагами доминируют высшие вегетативные центры, которые не являются симпатическими или парасимпатическими, а объединяют в себе регуляцию обоих отделов вегетативной нервной системы. К ним относится и ретикулярная формация. Они являются надсегментарными и расположены в стволе и плаще мозга, а именно: 1. Задний мозг: сосудодвигательный центр на дне IV желудочка; мозже 2. Средний мозг: серое вещество водопровода. 3. Промежуточный мозг: hypothalamus (tuber cinereum). 4. Конечный мозг: кора полушарий большого мозга. Наибольшее значение для вегетативной регуляции имеет гипоталамическая область, которая является одним из самых древних отделов головного мозга, хотя и в ней различают более старые "образования и филогенетически более молодые. Гипоталамо-гипофизарная система, действуя с помощью инкретов гипофиза, является регулятором всех эндокринных желез. Гипоталамическая область регулирует деятельность всех органов растительной жизни, объединяя и координируя их функции. Объединение вегетативных и анимальных функций всего организма осуществляется в коре большого мозга, особенно в премоторной зоне. Кора, будучи, по И. П. Павлову, комплексом корковых концов анализаторов, получает раздражения от всех органов, в том числе и от органов растительной жизни, и через посредство своих эфферентных систем, в том числе и вегетативной нервной системы, оказывает влияние на эти органы. Следовательно, существует двусторонняя связь коры и внутренностей — кортиковисцеральная связь. Благодаря этому все вегетативные функции подчиняются коре головного мозга, которая ведает всеми процессами организма. Таким образом, вегетативная нервная система есть не автономная система, как это считали до И. П. Павлова, а специализированная часть единой нервной системы, подчиненная высшим отделам ее, включая и кору большого мозга. Поэтому, как и в анимальной нервной системе, в вегетативной можно различать центральный и периферический ее отделы. К центральному отделу относятся описанные выше очаги и центры в спинном и головном мозге, а к периферическому — нервные узлы, нервы, сплетения и периферические нервные окончания. В последнее время установлено, что вегетативные узлы имеют свою афферентную иннервацию, благодаря которой они находятся под контролем центральной нервной системы. Значительные отличия имеет рефлекторная дуга (рис. 341). Клеточное тело воспринимающего нейрона как для анимальной, так и для вегетативной нервной системы помещается в спинномозговом узле, ganglion spinale, куда стекаются афферентные пути как от органов животной жизни, так и от органов растительной жизни и который, таким образом, является смешанным анимально-вегетативным узлом. Клеточное тело вставочного нейрона вегетативной нервной системы в отличие от анимальной нервной системы помещается в боковых рогах спинного мозга. При этом аксон вставочного анимального нейрона, исходящий из клеток заднего рога, заканчивается в пределах спинного мозга среди клеток его передних рогов. Что же касается вставочного нейрона вегетативной нервной системы, то он в спинном мозге не заканчивается, а выходит за его пределы, к нервным узлам, расположенным на периферии. Выйдя из спинного мозга, аксон вставочного нейрона подходит или к узлам симпатического ствола, ganglia trunci sympathici, относящимся к симпатическому отделу вегетативной нервной системы (они образуют симпатический ствол), или волокна не заканчиваются в этих узлах, а направляются к предпозвоночным узлам, расположенным более периферично, между симпатическим стволом и органом (ganglia coeliaca, ganglia mesenterica). Эти узлы также относятся к симпатической системе. Наконец, волокна могут доходить, не прерываясь, до узлов, лежащих или около органа {околоорганные узлы, например ganglia ciliare, oticum и др.), или в толще органа (внутриорганные, интрамуралъ-ные узлы); и те и дугие называют конечными узлами (ganliga terminalia). Они относятся к парасимпатическому отделу вегетативной нервной системы. Кроме макроскопически видимых обособленных узлов, по ходу вегетативных нервов встречаются мигрировавшие сюда в ходе эмбрионального развития небольшие группы эффекторных нейронов — микроганглии. Все волокна, идущие до узлов первого, второго или третьего порядка и являющиеся аксонами промежуточного нейрона, называются предузловыми волокнами, rami preganglionares. Они покрыты миелином. Третий, эффекторный, нейрон анимальной рефлекторной дуги помещается в передних рогах спинного мозга, а эффекторный нейрон вегетативной рефлекторной дуги вынесен в процессе развития из центральной нервной системы в периферическую, ближе к рабочему органу, и располагается в вегетативных нервных узлах. Из такого расположения эффекторных нейронов на периферии вытекает главный признак вегетативной нервной системы — двухнейронность эфферентного периферического пути: первый нейрон — вставочный; тело его лежит в вегетативных ядрах черепных нервов или боковых рогах спинного мозга, а нейрит идет к узлу; второй — эфферентный, тело которого лежит в узле, а нейрит достигает рабочего органа. Эффектор-ные нейроны симпатических нервов начинаются в ganglia trunci sympathici (узлы первого порядка) или ganglia intermedia (узлы второго порядка), а для парасимпатических нервов — в около- или внутриорганных узлах, ganglia terminalia (третьего порядка); так как в названных узлах осуществляется связь вставочных и эфферентных нейронов, то отмеченная разница между симпатическим и парасимпатическим отделами вегетативной нервной системы связана именно с этими нейронами. Аксоны эфферентных вегетативных нейронов почти лишены миелина — безмиелиновые (серые). Они составляют послеузловые волокна, rami postgang-lionares. Послеузловые волокна симпатической нервной системы, отходящие от узлов симпатического ствола, расходятся в двух направлениях. Одни волокна идут к внутренностям и составляют висцеральную часть симпатической системы. Другие волокна образуют rami communicantes grisei, соединяющие симпатический ствол с анимальными нервами. В составе последних волокна достигают соматических органов (аппарата движения и кожи), в которых иннервируют непроизвольную мускулатуру сосудов и кожи, а также железы. Совокупность описанных эфферентных вегетативных волокон, идущих от узлов симпатического ствола до органов сомы, составляет соматическую часть симпатического отдела. Такая структура обеспечивает функцию вегетативной нервной системы, которая регулирует обмен веществ всех частей организма применительно к непрерывно изменяющимся условиям среды и условиям функционирования (работы) тех или иных органов и тканей. Соответственно этой наиболее универсальной своей функции, связанной не с какими-либо отдельными органами и системами, а со всеми частями, со всеми органами и тканями организма, вегетативная нервная система и морфологически характеризуется универсальным, повсеместным распространением в организме. Следовательно, симпатический отдел иннервирует не только внутренности, но и сому, обеспечивая в ней обменные и трофические процессы. В результате каждый орган, по И. П. Павлову, находится под тройным нервным контролем, в связи с чем он различает три вида нервов: 1) функциональные, осуществляющие функцию данного органа; 2) сосудодвигательные, обеспечивающие доставку крови к органу, и 3) трофические, регулирующие усвоение из доставленной крови питательных веществ. Висцеральная часть симпатического отдела содержит все эти три вида нервов для внутренностей, а соматическая часть — только сосудодвигательные и трофические. Что же касается функциональных нервов для органов сомы (скелетная мускулатура и др.), то они идут в составе соматической, ани-мальной, нервной системы. Таким образом, основное отличие эфферентной части вегетативной нервной системы от эфферентной части анимальной заключается в том, что анимальные, соматические, нервные волокна, выйдя из центральной нервной системы, идут до рабочего органа, нигде не прерываясь, тогда как вегетативные волокна на своем пути от мозга до рабочего органа прерываются в одном из узлов первого, второго или третьего порядка. Вследствие этого эфферентный путь вегетативной нервной системы разбивается на две части, из которых он и состоит: предузловые миелиновые волокна, rami preganglionares, и послеузловые, лишенные миелина (безмиелиновые) волокна, rami postganglionares. Наличие узлов в эфферентной части рефлекторной дуги составляет характерный признак вегетативной нервной системы, отличающий ее от анимальной (см. рис. 341). Определенные отличия имеют и нервы. Афферентные пути вегетативной нервной системы не имеют характера макроскопически видимых нервов, а их волокна идут в составе других нервов (nn. splanchnici major et minor, n. vagus, задние корешки и др.). При этом для симпатического отдела характерно то, что связанная с ним чувствительная иннервация может распространяться на значительные расстояния и, следовательно, симпатический отдел может рассматриваться как система окольной иннервации. Так, например, афферентные спинальные нервные волокна, участвующие в формировании чревного сплетения, иннервирующего органы брюшной полости, происходят из многочисленных спинномозговых узлов (Cv — Lm). Это обстоятельство определяет множественность и многосегментар-ность путей и источников афферентной иннервации органов брюшной полости. Этим же объясняется и то, что чувство боли от внутренностей может передаваться как по вегетативным, так и по анимальным нервам. Существуют также собственные афферентные нейроны вегетативной нервной системы, замыкающиеся в вегетативных ганглиях, которые могут рассматриваться как периферические центры. Наряду с этим имеется не только диффузность в отношении распределения чувствительных нейронов и их волокон, но и преимущественное участие определенных спинномозговых узлов в иннервации внутренностей. Следовательно, среди источников и путей афферентной иннервации внутренностей можно выделить основные и дополнительные. Это деление тесно связано с представлением об окольных путях афферентной спинальной иннервации внутренностей. Окольные пути в патологических условиях (перерыв спинного мозга и др.) могут играть роль компенсаторных путей, возмещающих функцию нарушенных основных путей, компенсаторных приспособлений в виде «перекрытия» в афферентной иннервации органов. Что же касается эфферентных путей вегетативной нервной системы, то они образуют ясно выраженные нервы и узлы. Поэтому можно говорить о двух центробежных путях единой нервной системы: один путь — это анимальные, соматические, двигательные нервы, а другой — вегетативные. Вегетативные нервы образуют сплетения вокруг кровеносных сосудов, вместе с которыми они подходят и входят в органы. Наличие сплетений вокруг сосудов составляет характерный признак вегетативной нервной системы, отличающий ее от анимальной. Как уже отмечалось, вегетативная нервная система характеризуется универсальным, повсеместным распространением в организме. Она имеет широкую область эфферентной иннервации, охватывающую все органы и ткани тела, не исключая и скелетной мускулатуры (последнюю она тонизирует). В этом и сортоит морфологическая особенность вегетативной нервной системы в противоположность анимальной, которая иннервирует центробежными волокнами только скелетные мышцы, т. е. имеет сравнительно ограниченную область эфферентной иннервации. Для понимания строения необходимо учитывать развитие вегетативной нервной системы. Гладкая мускулатура беспозвоночных регулируется ганглиозно-сетевидной нервной системой, которая, кроме этой специальной функции, регулирует также и обмен веществ. Приспособление уровня обмена веществ к изменяющейся функции органов называется адаптацией (adaptare — прилаживать), а соответственная функция нервной системы — адаптационно-трофической (Л. А. Орбели). Адаптационно-трофическая функция есть наиболее общая и весьма древняя функция нервной системы, существовавшая у примитивных предков позвоночных. В дальнейшем ходе эволюции сильнее всего прогрессировали аппарат движения (развитие твердого скелета и скелетной мускулатуры) и органы чувств, т. е. органы животной жизни. Поэтому та часть нервной системы, которая была связана с ними, т. е. анимальная часть нервной системы, претерпела наиболее резкие изменения и приобрела новые признаки, в частности: изоляция волокон при помощи миелиновых оболочек, большая скорость проведения в'озбуждения (100—120 м/с). Напротив, органы растительной жизни претерпели более медленную и менее прогрессивную эволюцию, поэтому связанная с ними часть нервной системы сохранила за собой наиболее общую функцию — адаптационно-трофическую. Эта часть нервной системы и еств вегетативная- нервная система. Наряду с некоторой специализацией она сохранила ряд древних примитивных черт: отсутствие у большинства нервных волокон миелиновых оболочек (безмиелиновые волокна), меньшая скорость проведения возбуждения (0,3 — 10 м/с), а также меньшая концентрация и централизация эффекторных нейронов, оставшихся разбросанными на периферии, в составе ганглиев, нервов и сплетений. При этом эффекторный нейрон оказался расположенным вблизи рабочего органа или даже в толще его. Такое периферическое расположение эффекторного нейрона обусловило главную морфологическую особенность вегетативной нервной системы — двухнейронность эфферентного периферического пути, состоящего из вставочного и эффекторного нейронов. С появлением туловищного мозга (у бесчерепных) возникающие в нем импульсы адаптации идут по вставочным нейронам, обладающим большей скоростью возбуждения; выполняется же адаптация непроизвольной мускулатурой и железами, к которым подходят эффекторные нейроны, отличающиеся медленной проводимостью. Это противоречие разрешается в процессе эволюции благодаря развитию специальных нервных узлов, в которых устанавливаются контакты вставочных нейронов с эффекторными, причем один вставочный нейрон вступает в связь со многими эффекторными (примерно 1:32). Этим достигается переключение импульсов с миелиновых волокон, обладающих большой скоростью проведения раздражений, на безмиелиновые, обладающие малой скоростью. В результате весь эфферентный периферический путь вегетативной нервной системы разбивается на две части — предуз-ловую и послеузловую, а сами узлы становятся трансформаторами темпов возбуждения с быстрых на медленные. У низших рыб, когда образуется головной мозг, в нем развиваются центры, объединяющие деятельность органов, вырабатывающих внутреннюю среду организма. Так как в этой деятельности, кроме гладкой мускулатуры, принимает участие и скелетная (исчерченная), то возникает потребность в координации работы гладких и поперечнополосатых мышц. Например, жаберные крышки приводятся в движение скелетной мускулатурой, так же и у человека в акте дыхания участвует как гладкая мускулатура бронхов, так и скелетные мышцы грудной клетки. Такую координацию осуществляет развивающийся в заднем мозге специальный рефлекторный аппарат в виде системы блуждающего нерва (бульбарный отдел парасимпатической части вегетативной нервной системы). В центральной нервной системе возникаюх и другие образования, которые подобно блуждающему нерву выполняют функцию координации совместной деятельности скелетной мускулатуры, обладающей быстрой скоростью возбуждения, и гладкой мускулатуры и желез, обладающих медленной скоростью. Сюда относится та часть глазодвигательного нерва, которая осуществляет при помощи исчерченных и неисчерченных мышц глаза стандартную установку ширины зрачка, аккомодации и конвергенции соответственно силе освещения и расстоянию до рассматриваемого объекта по тем же принципам, как это делает фотограф (мезэнцефалический отдел парасимпатической части вегетативной нервной системы). Сюда относится и та часть крестцовых нервов (И —IV), которые осуществляют стандартную функцию тазовых органов (мочевого пузыря и прямой кишки) — опорожнение, в которой участвуют каж непроизвольные мышцы этих органов, так и произвольные мышцы таза и брюшного пресса — сакральный отдел парасимпатической части вегетативной нервной системы. В среднем и промежуточном мозге развился центральный адаптационный аппарат в виде серого вещества вокруг водопровода и серого бугра (hypothalamus). Наконец, в коре мозга возникли центры, объединяющие высшие анимальные и вегетативные функции. Развитие вегетативной нервной системы в онтогенезе (эмбриогенезе) идет иначе, чем в филогенезе. Вегетативная нервная система возникает из общего с анимальной частью источника — нейроэктодермы, чем доказывается единство всей нервной системы. Из общего зачатка нервной системы выселяются симпатобласты, которые скапливаются в определенных местах, образуя сначала узлы симпатического ствола, а затем промежуточные узлы, а также нервные сплетения. Отростки клеток симпатического ствола, объединяясь в пучки, образуют rami communicantes grisei. Сходным образом развивается и часть вегетативной нервной системы в области головы. Зачатки парасимпатических узлов выселяются из продолговатого мозга или ганглиоз-ной пластинки и совершают дальнюю миграцию вдоль ветвей тройничного, блуждающего и других нервов, оседая по их ходу или образуя интрамуральные ганглии. Дата добавления: 2015-12-16 | Просмотры: 1010 | Нарушение авторских прав |