|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
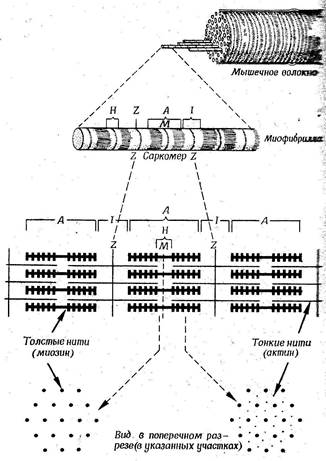
Скоростно-силовых качествСкелетные, гладкие и сердечная мышцы относятся к категории эффекторов – исполнительных органов. Сокращением и расслаблением скелетных мышц управляет сознание человека, а их тонус регулируется без его участия. Управление скелетными мышцами осуществляется мотонейронами, входящими в соматическую нервную систему и их сокращение происходит под воздействием нервных импульсов. Мышечные волокна сердца не контролируются волей, сознанием человека и управляются симпатическими и блуждающими нервами, являющимися компонентами вегетативной нервной системы (ВНС). Гладкие мышцы являются составной частью стенок кровеносных сосудов и полых внутренних органов. Они управляются вегетативной нервной системой и почти не подчиняются сознанию. Сократительная функция мышц обеспечивает силу, скорость, мощность процесса движения. Параметры этих двигательных качеств зависят от морфологической структуры мышц, особенностей течения комплекса биохимических процессов, обеспечивающих их сокращение и расслабление, а также зависят от качества различных регуляторных процессов и воздействий. Ультраструктура с келетных мышц. Они состоят из пучков мышечных волокон, число которых может достигать нескольких тысяч. И чем больше этих волокон, тем выше показатели двигательных качеств. Вся мышца, пучки волокон и отдельные волокна покрыты соединительнотканной оболочкой, в которой проходят сосуды и нервы. Мышечное волокно представляет собой громадную многоядерную клетку, длина которой может достигать 10-50см, при толщине не более 0,1мм. К волокну подходят окончания двигательного нерва, который может иннервировать группу мышечных волокон, именуемую двигательной (нейромоторной) единицей. Сила и скорость сокращения мышцы зависят от количества участвующих в сокращении двигательных единиц и от частоты нервной импульсации. Электровозбудимая оболочка (мембрана) мышечной клетки называется сарколеммой, ее цитоплазма – саркоплазмой. К сарколемме подходят окончания двигательных нервов, образуя синапсы (нервно-мышечное соединение). Основное пространство клетки занимают миофибриллы (волоконца), которых может быть более тысячи. По периферии клетки расположены многочисленные ядра. От сарколеммы внутрь отходят Т-трубочки, рядом с которыми находятся многочисленные «цистерны» саркоплазматического ретикулума, образующие единую систему сообщающихся цистерн. При микроскопическом исследовании мышц выявляется регулярная поперечная исчерченность, которая обусловлена наличием темных и светлых зон (дисков) в миофибриллах. В середине каждого светлого диска присутствует тонкая темная линия, именуемая Z-мембраной. Фрагмент миофибриллы между двумя соседними Z-мембранами называется саркомером. Пространство саркомера заполнено нитями (миофиламентами) двух сократительных белков – актина (тонкие нити) и миозина (толстые нити). Миозин, кроме того, является ферментом, катализирующим расщепление АТФ. Актиновые нити, расположенные эксцентрично, одним концом прикреплены к Z-мембране. Миозин и актин обуславливают поперечнополосатую исчерченность мышц и наличие (соответствено) темных и светлых дисков. Молекулы миозина представляют собой длинные асимметричные нити с многочисленными боковыми ответвлениями в форме шарообразной головы. Молекула толстой нити состоит из двух, скрученных между собой (двуспиральная структура), необычайно длинных, полипептидных цепей, каждая из которых содержит около 1800 аминокислотных остатков. Сокращение мышц происходит за счет образования боковыми ответвлениями миозина соединений (поперечных мостиков, актомиозиновых комплексов) между нитями актина и миозина и втягивания актиновых нитей в систему миозиновых нитей.
Рис 1.3.1. Строение мышцы
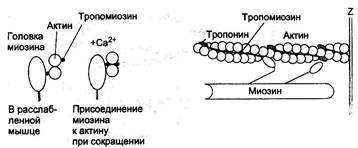
Миозиновые нити, расположенные в саркомере центрально, на ¼ своей длины, входят в промежутки между параллельно расположенными актиновыми нитями. При максимальном сокращении мышцы саркомеры укорачиваются на 20-50 %. Пассивное растяжение может удлинять ее до 120 %. Длина саркомера при сокращении уменьшается, но не меняется размер миозиновых и актиновых нитей. Актиновые нити образованы двойной, скрученной цепью глобулярных белковых молекул-мономеров (рис. 1.3.2.) и, в свою очередь, они оплетены тонкими нитями другого белка (фибриллярного) – тропомиозина. С тропомиозиновыми нитями соединяются молекулы еще одного белка – тропонина (глобулярный белок), который, как и тропомиозин, участвует в регуляции взаимодействий между актином и миозином (их соединение и разъединение) при сокращении и расслаблении мышц.
Рис. 1.3.2. Сократительные и регуляторные белки миофибрилл
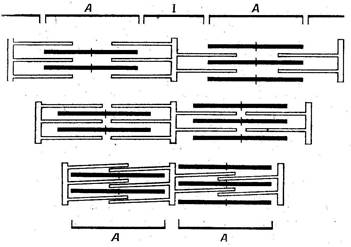
В состоянии покоя миозиновые и актиновые нити не могут взаимодействовать между собой, в связи с блокирующим влиянием тропонина и тропомиозина. Сигналом для сокращения мышц служит электрический импульс, приходящий из двигательного нерва и быстро распространяющийся по всей сарколемме. Происходит деполяризация мембраны, которая становится проницаемой для катионов К+, Na+, Са2+. Возбуждение сарколеммы вызывает выход Са2+ из цистерн саркоплазматической сети в саркоплазму. Это инициирует взаимодействие АТФ с толстыми и тонкими нитями. При этом ионы Са2+ выполняют еще одну функцию - стимулируют гидролиз АТФ миозином (миозиновой АТФ-азой). Связывание кальция (Са-2+) с тропонином вызывает структурное изменение тонкого филамента – участок присоединения на актине оказывается открытым, и миозин с актином получают возможность взаимодействовать между собой. Так, с помощью шарообразной головки миозина образуется “поперечный мостик” (“спайка”) между миозином и актином. Мышечное сокращение является результатом превращения химической энергии, аккумулированной в макроэргических связях молекулы АТФ, в механическую работу. Точный механизм этого превращения неизвестен. Предполагается, что распад АТФ сопровождается освобождением энергии необходимой для вращательного движения поперечного мостика. Это движение приводит к скольжению и втягиванию актиновых нитей между миозиновыми. Движение происходит в каждом саркомере миофибриллы. Возникающая сила сокращения мышцы обусловлена цикличностью образования и разрушения множества поперечных мостиков между миозином и актином. Миозиновые головки одновременно присоединяются к нитям актина, соединенным с двумя соседними Z – мембранами и тянущее усилие сближает эти мембраны, уменьшая длину саркомера. Высокая концентрация кальция в саркоплазме активирует обратное движение этих ионов в саркоплазматический ретикулум (эффект “кальциевого насоса”), используя для этого энергию АТФ. Тропонин, освободившись от кальция, восстанавливает свое блокирующее влияние, а также исходную конформацию молекулы, головки миозиновых нитей не могут взаимодействовать с актином и образовывать поперечные мостики. Так наступает расслабление мышечного волокна. К настоящему времени накоплено достаточно данных, свидетельствующих о генетической обусловленности биохимических и физиологических процессов, обеспечивающих врожденную уникальную двигательную одаренность некоторых спортсменов. Установлено, что у животных и человека имеются три вида мышечных волокон – белые, красные и промежуточные. Белые мышечные волокна входят в состав тетанических нейромоторных единиц (быстрый, утомляемый тип), обеспечивающих, прежде всего, быстрые движения. Они лишены миоглобина, содержат много креатинфосфата, гликогена и обладают большими гликолитическими возможностями, высокой активностью ферментов. В белых волокнах больше сократительного белка миозина, являющегося одновременно ферментом (аденозинтрифосфатазой), расщепляющим АТФ и обеспечивающим, в значительной степени, “взрывную силу” и “стартовую скорость”. Рис. 1.3.3. Модель сокращения мышцы
В быстро сокращающихся белых волокнах значения максимальной скорости их сокращения почти в 4 раза выше, чем в медленно сокращающихся красных мышечных волокнах. Высокую стартовую скорость обеспечивают так же возрастающее в процессе тренировок количество ферментов, катализирующих расщепление аденозинтрифосфата (миозиновая АТФ-аза) и креатинфосфокиназную реакцию. Увеличение количества ферментов ведет к возрастанию их активности, кроме того, совершенствуется химическая структура этих биологических катализаторов, делая ферменты еще более активными. Известно так же, что диаметр нервных волокон, подходящих к белым мышечным волокнам, больше. Это благоприятствует движению большего потока нервных импульсов, идущих из ЦНС к нейромоторной единице и достигающих синапсов на наружной мембране мышечного волокна. Большое значение имеют синаптогенез (увеличение количества синапсов), степень разветвленности синапсов и количество образующегося в них нейромедиатора ацетилхолина. Скоростно-силовые качества связаны с эффективностью работы эндокринной системы, выбросом адреналина, глюкагона, со скоростью передачи электрического возбуждения от наружной мембраны к миофибриллам, величиной потока ионов Са-2+, освобождающихся из внутренних цистерн саркоплазматического ретикулума во внутриклеточное пространство, скоростью развития активации в миофибриллах. Эти качества зависят от особенностей строения сократительных белков миофибрилл (миозина, актина), их массы, количества калия, натрия и магния в клетках. У “прирожденных” спринтеров масса белых мышечных волокон больше, чем красных. Максимальная мышечная сила пропорциональна длине саркомера, или длине миозиновых (толстых) нитей и общему содержанию в мышце сократительного белка актина. От этого зависит количество спаек образующихся между актином и миозином, обуславливающих скорость и мощность мышечного сокращения. Если длина саркомера, степень полимеризации миозина – это неизменный, генетически обусловленный фактор, который не поддается влиянию тренировки, то содержание миозина и актина в клетках возрастает под влиянием тренировок. Красные мышечные волокна входят в состав тонических нейромоторных единиц (медленный, малоутомляемый тип). В них много миоглобина и митохондрий, обеспечивающих эффективное аэробное энергообразование. В этих волокнах существенно меньше креатинфосфата, гликогена, миозина и, следовательно, меньше анаэробный потенциал энергообразования. Тонические нейромоторные единицы обеспечивают длительные движения, но невысокой интенсивности – длительную работу умеренной и большой мощности. Существует также промежуточный (быстрый, малоутомляемый) тип двигательных единиц. Таким образом, мышечная система спортсмена состоит из перечисленных типов нейромоторных систем, развитие которых у разных людей различно в силу генетических причин. Это имеет большое значение в спортивной практике, так как скоростно-силовые возможности определяются количеством белых мышечных волокон у индивида. Биопсия мышц позволяет, на этапе раннего отбора, выявлять потенциальных спринтеров. Движения человека оцениваются также величиной мощности развиваемого усилия, которая является произведением силы на скорость. Значения максимальной мощности существенно изменяются в процессе адаптации к специфическому виду двигательной деятельности. Так в быстросокращающихся волокнах мощность достигает 155 Вт. кг -1 массы мышц, а в медленносокращающихся волокнах – 40 Вт.кг-1. Соответственно, в икроножной мышце спринтера содержание быстросокращающихся волокон достигает 60 %, а у стайеров таких волокон только 35 %. И, напротив, у стайеров в икроножных мышцах на медленносокращающиеся волокна приходится более 80 %, а у спринтеров – только 23 % (Н.И. Волков с соавт., 2000). Проявления скоростно-силовых качеств у спортсмена связаны с координацией работы мышц-синергистов и мышц-антагонистов, с особенностями строения этих мышц. В частности, они зависят от направления сухожильных тяжей и расположения относительно них мышечных волокон. От этого зависит величина суммарного мышечного усилия, развиваемого в точках прикрепления сухожильных окончаний мышц к костным рычагам.
Дата добавления: 2015-12-16 | Просмотры: 666 | Нарушение авторских прав |