|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Айала Ф., Кайгер Дж. Современная генетика: В 3-х т. Т. 3. Пер. с англ.: – М.: Мир, 1988. – 336 с.228 Эволюция генетического материала
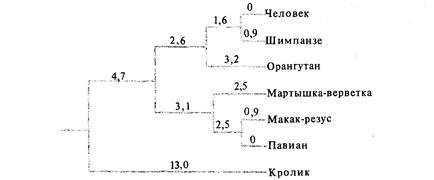
люционный консерватизм цитохрома с позволяет использовать его для анализа генетических различий между организмами, находящимися лишь в отдаленном родстве. Однако в силу того же эволюционного консерватизма цитохром с оказывается бесполезным при исследовании эволюционных изменений у близкородственных организмов, так как у них молекулы цитохрома с полностью или почти полностью идентичны. Например, у человека и шимпанзе первичная структура цитохрома с совершенно одинакова, хотя пути их эволюции разошлись 10-15 млн. лет назад. Цитохромы с человека и макака-резуса различаются только по одной аминокислоте, хотя общий предок этих организмов существовал 40-50 млн. лет назад. Для разных белков характерны различные скорости эволюции. При анализе филогенетических различий между близкородственными организмами можно использовать аминокислотные последовательности быстро эволюционирующих белков, таких, как фибринопептиды млекопитающих (рис. 26.11). Карбоангидразы - это быстро эволюционирующие белки, играющие важную физиологическую роль при обратимой гидратации СО2, а также в некоторых секреторных процессах. На рис. 26.12 изображено филогенетическое древо некоторых приматов, построенное на основе данных об аминокислотной последовательности карбоангидразы I с указанием минимально необходимого числа нуклеотидных замен в каждой ветви древа. Генетические изменения, происходящие в ходе эволюции близкородственных видов, можно изучать также с помощью других методов, таких, как гибридизация ДНК, электрофорез в гелях и иммунологические методы. Дата добавления: 2015-12-16 | Просмотры: 531 | Нарушение авторских прав |