|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Айала Ф., Кайгер Дж. Современная генетика: В 3-х т. Т. 3. Пер. с англ.: – М.: Мир, 1988. – 336 с.26. Видообразование и макроэволюция 237
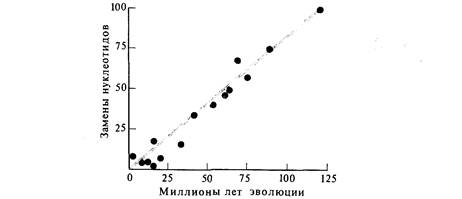
тельно скорости эволюции другого остается постоянной во времени. Правдоподобность этой гипотезы лежит на границе достоверности (вероятность χ 0,06). Вероятность того, что все наблюдаемые изменения обусловлены случайными причинами, крайне мала и составляет 6 · 10–б. Полученные таким образом результаты имеют особую ценность, поскольку в работе не использовалось никаких палеонтологических датировок. Филогения строилась исключительно на основе данных о белках. Это максимизирует вероятность соответствия между использованными данными и гипотезой о том, что скорость молекулярной эволюции была стохастически постоянной. Несмотря на это, указанные данные не подтверждают гипотезы о пуассоновском характере процесса молекулярных изменений, т.е. о постоянстве их вероятности. Однако недавно Джон Гиллеспай и Чарлз Лэнгли показали, что данные, представленные в табл. 26.9, удовлетворяют гипотезе о постоянстве скорости молекулярной эволюции, если отказаться от предположения о пуассоновском характере процесса и допустить, что варианса процесса больше, чем для соответствующего пуассоновского распределения. Независимо от того, является ли скорость молекулярной эволюции стохастически постоянной, ясно, что разброс значений скорости эволюции не очень велик. Это значит, что генетические данные можно использовать в качестве эволюционных часов, хотя и не очень точных. Для то- Дата добавления: 2015-12-16 | Просмотры: 571 | Нарушение авторских прав |