|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Биохимические. (Ниренберг – бесклеточные системы)1. Добавив синтетическую полиуридиловую РНК (полиU) в бесклет. сист., приготовленную из E.coli, обнаружили, что полиU стимулирует включение в полипептид только одного типа АК ост. – Фен. Таким обр., учитывая триплетность кода, кодон для Фен был расшифрован как UUU в иРНК. Стали исп. этот способ для разных нуклеотидов, выяснили состав, но не последовательность. 2. Г. Коран: метод хим. синтеза ДНК-подобных полимеров с заданной посл-ю нуклеотидов – матрица для синтеза РНК – при помощи ДНК-завис. РНК-полимеразы пол. РНК с заранее известной посл-ю и исп. в бескл. сист. синтеза белка. 3. Ниренберг и Ледер: исп. то, что промеж. пр-ты в синтезе белка – АК, связ. с тРНК. Одного тринуклеотида на рибосоме достаточно для связывания и с рибосомой, и с тРНК.
47. Свойства нуклеиновых кислот, определяющиее их генетические функции. информативность - специфическая последовательность четырех нуклеотидов в полинуклеотидной цепи точность полуконсервативный механизм репликации ДНК стабильность – двуспиральная структура, способность к самовоспроизведению (репликации) пластичность мутационная изменчивость -> новые признаки рекомбинационная изменчивость - механизмы обмена генетическим материалом, принадлежащим разным особям, "по горизонтали", и возникновение на базе этого особей с рекомбинантным геномом. При генетических рекомбинациях новых генов в генофонде, как правило, не появляется. В результате ген рекомб обеспечивается возможность объединения разных генов и создание разных вариантов генных сочетаний в геноме видовая специфичность состава ДНК (нач 50-х гг) Э. Чаргафф показал, что соотношения 4 сортов ее мономеров - G, А, С и Т - различ у разн видов орг-мов
Репликация ДНК Принципы репликации Комплементарность. Каждая из двух цепей "материнской" молекулы ДНК служит матрицей для синтеза дополняющей её, т.е. комплементарной "дочерней" цепи Полуконсервативность. В результате репликации образуются две двойные "дочерние" спирали, каждая из которых сохраняет (консервирует) в неизменном виде одну из половин "материнской" ДНК. Антипараллельность и униполярность. Две комплементарные цепи в молекуле ДНК ориентированы в противоположных направлениях - антипараллельно. При этом синтез комплементарных нитей всегда ведётся в 3' направлении, то есть униполярно. Поэтому в процессе репликации одновременный синтез новых цепей идёт антипараллельно Прерывистость. репликация начинается одновременно в нескольких местах молекулы ДНК. Участок между двумя точками, в которых начинается синтез "дочерних" цепей, называется репликоном. В эукариотической клетке в каждой молекуле ДНК имеются в зависимости от размеров сотни и даже тысячи репликонов, в которых можно видеть "репликативную вилку". Ведут синтез два по разному организованных репликативных ферментативных комплекса. Одна "дочерняя" цепь (лидирующая) растёт непрерывно, а другая (отстающая) - в виде " фрагментов Оказаки ". Механизм синтеза "дочерних" цепей ДНК фрагментами называют прерывистым. Потребность в затравке. Его синтезирует особая РНК-полимераза, называемая праймазой.
48. Генетический контроль и энзимология процесса репликации ДНК. События в репликативной вилке. Ошибки репликации возникают из-за возможности существования оснований ДНК в неск таутомерных формах. Спонтанная мутабильность повышается в результате мутационного изменения генов, контролирующих репликацию ДНК. Ген-мутатор: мутация гена 43 (контролир репликативн ДНК-полимеразу, которая обладает двумя активностями: 5-3-полимеразной и 3-5-экзонуклеазной(последняя – функции коррекции при репликации)) бактериофага Т4 (Дж Спейсер, 1965г) -> повыш частота мутаций в гене rII, контролир скорость лизиса. Ген-антимутатор: также мутация гена 43 фага Т4 -> понижен мутагенез под действием 2-аминопурина: антимутаторная ДНК-полимераза включает меньше 2-аминоурина в ДНК, чем полимераза дикого типа или мутаторная. Т.о. частота ошибок репликации зависит от соотношения полимеразной и экзонуклеазной активности ДНК-полимеразы. Мутаторную и антимутаторную активность проявляют также и аллели других генов, контролирующих репликацию: гены, кодир ДНК-лигазу, ДНК-связ белки и др. Также есть данные о мутат активности гена dnaE, контролир ДНК-полимеразу у E.coli, а также обнаружение мутагенной ДНК-полимеразы в клетках человека, больного лейкемией. Мутаген, действующий непосредственно в репликативной вилке: N-метил-N-нитро-N-нитрозогуанидин (МННГ), который взаимодействует с одноцепочечными участками в вилке или действует на белки реплисомы. Процесс репликации ДНК согласован с клеточным делением и требует совместного действия многих белков. В нем участвуют: 1. ДНК-хеликаза и дестабилизирующие белки; они расплетают двойную спираль родительской ДНК и формируют репликационную вилку. 2. ДНК-полимеразы, которые катализируют синтез полинуклеотидной цепи ДНК в направлении 3'-5, копируя в репликационной вилке матрицу с высокой степенью точности. Поскольку две цепи двойной спирали ДНК антипараллельны, в направлении 5'-3' непрерывно синтезируется лишь одна из двух цепей, ведущая; другая цепь, отстающая, синтезируется в виде коротких фрагментов Оказаки. ДНК-полимераза способна к исправлению собственных ошибок, но не может самостоятельно начать синтез новой цепи. 3. ДНК-праймаза, которая катализирует короткие молекулы РНК-затравки. Впоследствии фрагменты РНК удаляются - их заменяет ДНК. 4. ДНК-топоизомеразы, помогающие решить проблемы кручения и спутывания спирали ДНК. 5. Инициаторные белки, связывающиеся в точке начала репликации и способствующие образованию нового репликационного глазка с одной или двумя вилками. В каждой из вилок вслед за инициаторными белками к расплетенной ДНК сначала присоединяется белковый комплекс, состоящий из ДНК-хеликазы и ДНК-праймазы (праймосома). Затем к праймосоме добавляются другие белки и возникает "репликационная машина", которая и осуществляет синтез ДНК. 6. РНК-полимеразы, кодир геном dna G – инциация репликации.
Спираль расплетается ДНК-хеликазой; этому процессу помогают ДНК-топоизомераза, раскручивающая цепи ДНК, и множество молекул дестабилизирующего белка, связывающихся с обеими одиночными цепями ДНК. В области вилки действуют две ДНК-полимеразы - на ведущей и отстающей цепи. На ведущей цепи ДНК-полимераза работает непрерывно, а на отстающей фермент время от времени прерывает и вновь возобновляет свою работу, используя короткие РНК-затравки, синтезируемые ДНК-праймазой. Молекула ДНК-праймазы непосредственно связана с ДНК-хеликазой, образуя структуру, называемую праймосомой. Праймосома движется в направлении раскрывания репликационной вилки и по ходу движения синтезирует РНК-затравку для фрагментов Оказаки. В этом же направлении движется ДНК-полимераза ведущей цепи и, хотя на первый взгляд это трудно представить, ДНК-полимераза отстающей цепи. Для этого последняя накладывает цепь ДНК, которая служит ей матрицей, саму на себя, что и обеспечивает разворот ДНК-полимеразы отстающей цепи на 180 градусов. Согласованное движение двух ДНК-полимераз обеспечивает координированную репликацию обеих нитей.
События в репликативной вилки Е. Coli: присоединение к хромосоме белка DnaA – разделение цепей и работа геликахы. Гираза решает топологические проблемы, связанные с разделением цепей двойной спирали. С образовавшейя одноцепочечной ДНК связываются белки SSB, стабилизирующие вилку репликации. Праймаза – синтезирует РНК-праймеры на лидирующей и отстающей цепях. У сайта начала реплиации много АТ пар. Инициация репликации у дрожжей: G1 – формирование пре-репликационного комплекса, в состав которого входят шесть белков комплекса ORC и белки Cdc6 и Mcm. При переходе к статдии S frlbdyjcnm Cdk1 возрастает и Cdc6p покидает комплекс, на его место встаёт Cdc45& После инициации репликации комплекс превращается в пост-репликационный, он состоит только из белков ORC, связв=анных с хроматином. Этот комплекс сохраняется до конца митоза, когда активность Cdk1 падает. Разделение двойной спирали происходит с помощью ДНК-геликазы и репликационного белка RPA. Репликационный белок A связывается с одноцепочечной ДНК, затем альфа-ДНК-полимераза-праймаза синтезирует короткие РНК-праймере на лидирующей и отстающей цепях, поссле чего происходит замена полимеразного комплекса на комплекс дельта-ДНК-полимеразы. Синтез новых цепей ДНК осуществляется ДНК-полимеразами. ДНК-лигазы сшивают фрагменты оказаки.
49.Повреждения днк Алкилирование (метилазе Dam), пиримидиновый димер и 6-4-фотопродукт (UV), одно- и двунитевой разрыв (γ-, X-rays), 8-оксигуанин (АФК), АП-сайт (гидролиз N-гликозидной связи), межнитевая сшивка (от митомицина C, азотистого иприта), сшивка ДНК-белок, ошибка спаривания (A-C, T-G и т. д.), дезаминирование оснований. Следствия: запуск системы репарации – эксцизионная репарация, репарация неспаренных оснований (Mismatch repair – MMR), пострепликативная рекомбинационная репарация. Если репарация не помогла, запускается SOS-репарация, которая ошибается. При низкой активности репаративных механизмов повреждения ДНК приводят к мутациям.
50. Мутагенез, вызванный химическими агентами аналогами оснований, интеркалирующими агентами, алкилирующими агентами, активными формами кислорода. Нарушения: ковалентные сшивки двух близко расположенных оснований, ковалетное связывание азотистых оснований с алифатическими и ароматическими радикалами; химические перестройки азотистых оснований, нарушения свхврофосфатного каркаса. Алкилирующие агенты – переносят алкилирующие группы на биологические макромолекулы. Этиленимины, алкилалкансульфаноаты, эпоксиды, многоатомные спирты, производные нитрозомочевины и аминоимидазола. Транзиции – основной тип изменений под действием алкилирущиах агентов. Аналоги онований – более высокая способность к таутомерным переходам, так же повышают чувствительность к другим мутагенам. Акридиновые красители – мутации типа рамки считывания.внедряются между соседними оснвоаниями ДНК – либо редупликация с новым основанием, либо ошибка в репарации и рекомбинации. Экзогенные ДНК – формирование множетства видимых и летальных перестроек – генные мутации, делеции. Очень пролонгированный эффект.
51. Мутагенез, вызванный физическими мутагенами – Ионизирующим излучением, УФ-излучением. Ионизирующее излучение – электромагнитные рентгеновские, гамма и космические лучи, а так же высокоэнергетические альфа, бета и другие корпускулярные излучния. Выбивают электроны из внешней оболочки атомом или молекул, превращая их в положительные ионы, освобожившиеся электрны выбивают электроны из других атомов и молекул.Аомы, захватившие такие электроны, приобретают -. Разрывается углеродно-фосфатный скелет ДНК, разрушаются основания (особенно пиримидиновые), происходят химические перестройки оснований, изменяющие способность к спариванию (например, производное пурина спар с пурином). Образуются сшивки как в молекулах нуклеиновых кислот, так и между молекулами ДНК и белками. Уф-лучине действуют на половые клетки, проникают в ткани очень слабо, не облаают достаточной энергией для индукции ионизации атомов и только возбудлают электронные оболочки. Сутации в клетках, образующих монослой. Возникают пиримидин-пиримидиновые димеры, так же под действием УФ-облучения эти димеры разрушаются. Присутствие диеров приводит к ошибкам при репликации.
52. Ошибки процесса репликации ДНК в клетках здорового человека происходят с частотой ~10-9-10-10 на 1 нуклеотид ДНК на 1 клеточное поколение. В каждой клетке при температуре тела человека от ДНК отрываются >10000 оснований (в основном пуриновых) вследствие разрывов N-гликозидных связей между основанием и дезоксирибозой. Частота спонтанных реакций дезаминирования цитозина в урацил – более 100 на клетку за сутки. В ходе репликации ДНК, транскрипции и других процессов постоянно рвутся цепи ДНК. И это только часть реакций деструкции ДНК, происходящих в нормальной клетке в обычных условиях. В то же время частота фиксированных наследственных изменений – мутаций генов несравнимо ниже частоты повреждений ДНК. Менее 1 повреждения ДНК из 1000 превращается в мутацию. Этот факт объясняется работой систем репарации повреждений в ДНК. Мутаторы – гены, повышающие частоту возникновения мутаций, антимутаторы – понижающие эту частоту (dam, mutH, mutL, mutS, mutU). Антимутаторными могут быть гены, кодирующие ферменты репликации, например, при мутации в полимеразном домене ДНК-полимераз. Мутаторными могут быть гены с мутацией в репаративном домене ДНК-полимеразы I.
Скорость репликации бактериальной хромосомы составляет около 1000 п.н./секунду. Скорость репликации у эукариот (млекопитающие) составляет около 100 п.н./с. Прежде, чем начать репликацию необходимо расплести ДНК. Это расплетание осуществляется специальным инициаторным белком DnaA. Сайты связывания этого белка находятся в ориджине. Кроме белка DnaA в раскрытии ДНК принимают участие и другие белки (FIS, HU, HIF). С помощью этих инициаторных белков на ДНК садится ДНК-хеликаза DnaB/DnaC (оба белка – гексамеры). Хеликаза двигается по ДНК и разрывает водородные связи между комплементарными основаниями, разделяет цепи и продвигает репликационную вилку. С однонитевыми участками ДНК связывается белок SSB. Это дестабилизирующий ДНК белок, он полностью покрывает однонитевые участки и предотвращает восстановление дуплекса. ДНК закручена в спираль, поэтому при продвижении репликационной вилки еще не удвоенная часть ДНК должна вращаться с большой скоростью. Эта топологическая проблема решается с помощью ДНК-топоизомераз, которые способны делать в цепях разрывы, менять скрученность ДНК, а потом зашивать эти разрывы, которые раскручивают ДНК впереди репликационной вилки. Топоизомераза I релаксирует суперскрученную ДНК, делая разрыв в одной цепи и протягивая вторую цепь через этот разрыв, и зашивает разрыв. Топоизомераза II, к которой относится ДНК-гираза, делает двунитевой разрыв. ДНК-полимеразы не могут начинать синтез ДНК на матрице, а способны лишь добавлять новые дезоксирибонуклеотиды к 3'- концу уже имеющейся цепи. Т.е. синтез ДНК на матрице должен быть инициирован коротким олигонуклеотидом, который формирует участок двунитевой ДНК. Такие короткие олигонуклеотиды называются затравками или праймерами. Праймаза (DnaG) образует комплекс с матричной ДНК и дополнительными белками, такими как DnaB, DnaT, PriA, PriB, PriC – это инициирующий комплекс или праймосома, которая синтезирует РНК-праймер. Все ДНК-полимеразы способны синтезировать новую цепь только в одном направлении- 5'-3'. Синтез ДНК осуществляется непрерывно только на одной нити, называемой ведущей (ДНК-полимераза III). На второй нити, называемой отстающей, ДНК синтезируется короткими (длиной от 100 до 1000 нуклеотидов) фрагментами, названными фрагментами Оказаки по имени обнаружившего их ученого. Таким образом, одна из нитей синтезируется в направлении, противоположном направлению передвижения вилки. Синтез фрагмента Оказаки терминируется перед началом РНК-праймера предыдущего фрагмента. Здесь начинает работать ДНК-полимераза I. Ее 5’-3’-экзонуклеазная активность удаляет РНК-затравку, и одновременно достраивает предыдущий фрагмент ДНК. Затем два состыкованных фрагмента Оказаки соединяются между собой ДНК-лигазой.
54. Репарация ДНК и мутационный процесс Мутации некоторых генов, ответственных за репарацию у Е. coli,бактериофага Т4, дрожжей, а также в клетках высших эукариот, проявляют мутаторный или антимутаторный эффект, подобно мутациям в генах, ответственных за репликативный комплекс. Наиболее подробно участие процессов репарации в возникновении мутаций исследовано у бактерии Е. coli. Показано, что мутация в гене uvr E, контролирующем ликвидацию однонитевых разрывов после ультрафиолетового (но не ионизирующего) облучения, повышает спонтанное возникновение транзиций AT - GC в 350—400 раз, а транзиций GC- АТ в 150—200 раз. Она повышает также частоту мутаций, индуцированных ультрафиолетовым светом и метилметансульфонатом. Изучение генетического контроля репарации (а также рекомбинации) позволило доказать участие некоторых нормальных процессов, происходящих в клетке, в превращении предмутационных изменений ДНК в мутации. В частности, оказалось, что процесс становления мутаций может быть генетически блокирован так же, как любой другой физиологический процесс. Так, изменение генов lex А или rеc А ведет к частичному или полному подавлению мутационного процесса под действием ультрафиолетового света, ионизирующих излучений и некоторых химических мутагенов. Э. Виткин обратила внимание на связь нескольких явлений, для которых общей причиной служит облучение клеток ультрафиолетовым светом: 1) индукция профага лямбда; 2) повышение выживаемости облученного бактериофага лямбда при заражении им предварительно облученных клеток Е. coli по сравнению с выживаемостью в необлученных клетках — так называемая W-реактивация, открытая Дж. Уэйглом; 3) блокирование клеточных делений у некоторых мутантов Е. coli, в результате чего клетки приобретают нитевидную форму; 4) повышение частоты рекомбинации; 5) повышение частоты мутаций. Оказалось, что W-реактивация бактериофага сопровождается повышением его мутабильности. Кроме того, явление W-реактива-ции так же зависит от дозы ультрафиолетового света, как и индукция профага лямбда. Для осуществления обоих процессов требуется нормальное состояние генов rec A+ и lex А +. У мутантов rес А и lex А они подавлены, как и образование нитей, не говоря уже о мутагенезе, индуцированном ультрафиолетовым светом. Параллелизм в проявлении индукции профага и W-реактивации (сопровождаемой повышением мутабильности) указывает на существование индуцируемой системы репарации, которая в связи с этим получила наименование SOS-репарации, т. е. репарации, включаемой для спасения. Индуцибельная система репарации действует по механизму пострепликативной репарации. На это указывает ее зависимость от гена rеc А. SOS-репарация включается в тех случаях, когда «безошибочная» дорепликативная система репарации не справляется с устранением повреждений или когда она блокирована мутационным путем. Для индукции системы SOS-репарации требуется 30—60 димеров тимина на геном Е. coli. Сигналом индукции служит задержка репликации, которая при этом наблюдается. Связь репарации и мутационного процесса показана для Drosophila melanogaster. Так, при действии кофеина на недокармливаемых мух наблюдается четкий антимутагенный эффект, судя по критерию спонтанных потерь Х-хромосомы. Известен мутатор ти у дрозофилы, в присутствии которого мухи проявляют повышенную чувствительность к рентгеновым лучам и метилметансульфонату. Мутатор ти находится в непосредственной близости к мутации С (3) G, блокирующей мейотическую рекомбинацию.
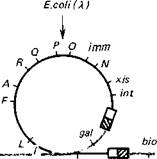
Рис. 9.8. Схема интеграции и эксцизии генома бактериофага γ за счет сайт-специфической рекомбинации с хромосомой Е. coli и образование трансдуцирующих частиц
вопрос 55 Mismatch repair – MMR. С неправильным основанием связывается белок MutS, с которым затем связываются белок MutL. Это событие активирует латентную эндонуклеазу MutH. Образуется репарационный комплекс с затратой 1 молекулы АТР. Белок MutH разрезает неметилированную нить ДНК по сайту GATC, который может располагаться по любую сторону от неправильного основания. Затем ДНК-хеликаза II (MutU = UvrD) расплетает надрезанную нить ДНК между надрезом и неспаренным основанием (включая его) и вытесняет ее из гетеродуплекса. Экзонуклеаза I (если это 3'-конец) или экзонуклеаза VII (если это 5'-конец) гидролизует вытесненную нить. Этот процесс нуждается в MutL и MutS. Вырезаются фрагменты порядка 1000 н. Затем образовавшаяся брешь застраивается ДНК-полимеразой III в присутствии белка SSB. В завершение, ДНК-лигаза восстанавливает фосфодиэфирную связь. Система MMR у эукариот организована сложнее функционирует эффективнее по сравнению с бактериями. У эукариот MMR исправляет все некомплементарные пары оснований и, кроме того, репарирует делеции или инсерции размером до 12 н. У бактерий MMR неспособна исправлять пары С*С и репарирует делеции/инсерции не более 3 н. Ключевые белки MMR – MutL и MutS высококонсервативны, их гомологи обнаружены у всех организмов от E.coli до человека.
56. Репарация ДНК Удивительная стабильность генетического материала — ДНК связана отнюдь не с ее консервативностью, а с существованием в клетках всех живых организмов специальных систем репарации, устраняющих из ДНК возникающие в ней повреждения. Явление репарации, или восстановления жизнеспособности клетки, после действия на нее гамма- и рентгеновых лучей было открыто в 1958 г. В. И. Корогодиным у диплоидных дрожжей. Повреждения ДНК, возникающие при действии излучений и химических агентов, в конечном счете приводят к нарушению регулярной Уотсон-Криковской структуры, что выражается в локальной денатурации молекулы и приводят к частичному или полному блокированию репликации. Именно такие нарушения конформации, а не конкретные изменения мономеров служат мишенью для большинства систем репарации ДНК. Существует несколько систем, осущ.репарацию. В каждую входит 4 типа ферментов:1й – узнающий «неправильный»участок и разрывающий цепь вблизи него, 2й – удал.повр.участок,3й-днк-полимераза,4й-днк-лигаза. В заав от типа повреждений меняются 1й и 2й фермент. У бактерий имеются по крайней мере 2 ферментные системы, ведущие Р. Первая осуществляет вырезание и ресинтез на небольшом участке в 5—7 нуклеотидов, вторая — на участке в тысячу нуклеотидов и более. Ферменты второй системы Р. участвуют также в процессах генетической рекомбинации. В случае повреждений, вызванных, например, УФ-светом, нормальная клетка кишечной палочки способна репарировать до 2000 повреждений; клетка с выведенной из строя первой системой Р. — около 100 повреждений; клетка с выведенными из строя обеими системами Р. погибает от одного повреждения. Существуют бактерии с исключительно активными ферментами Р. (например, Micrococcus radiodurans), которые благодаря этому способны выживать в воде, охлаждающей ядерные реакторы. Ферментные системы Р., как полагают, принимают участие и в нормальной репликации ДНК, т. е. её удвоении. При репликации материнская ДНК деспирализуется (раскручивается), что может сопровождаться разрывами её нитей. Кроме того, дочерние цепи ДНК синтезируются в виде небольших фрагментов. Поэтому заключительная фаза репликации — Р. всех дефектов, возникших при синтезе ДНК. Важная функция второй системы Р. — её участие в образовании мутаций. Под действием различных мутагенов в ДНК образуются производные нуклеотидов, чуждые клетке. Они устраняются системой Р., которая заменяет их на нуклеотиды, естественные для ДНК, но иногда измененные по сравнению с первоначальными. Открытие Р. ДНК привело к коренным изменениям представлений о молекулярных механизмах, обеспечивающих стабильность генетического аппарата клеток и контролирующих темп мутационного процесса. 57.Репарация ДНК –фотореактивация,деалкилирование,реп.однонитевых разрывов,р.АП-сайтов. Мутации, вызванные воздействием ультрафиолетового света, могут репарироваться благодаря явлению фотореактивации. Фотореактивация заключается в восстановлении биологической активности клеток или молекул ДНК, поврежденных ультрафиолетовым излучением в результате последующего воздействия видимого света. При фотореактивации происходит мономеризация циклобутановых димеров тимина и других пиримидиновых димеров in situ. Известна так называемая неферментативная коротковолновая фотореактивация, которая заключается в мономеризации димеров при действии ультрафиолетового света с длиной волны 240 нм, а также ферментативная фотореактивация. Именно последнюю обычно и подразумевают под собственно фотореактивацией. Фотореактивация при действии видимого света (300—400 нм — наиболее активная часть спектра) была обнаружена в 1949 г. в нескольких лабораториях. Механизм этого явления был раскрыт в начале 60-х годов нашего века после выделения К. Рупертом из клеток микроорганизмов фермента фотореактивации — дезоксирибопиримидинфотолиазы. Экстракты дрожжей оказались способными восстанавливать трансформирующую активность ДНК Haemophyllus influenzae на свету. Субстратом фермента фотореактивации служат димеры пиримидиновых оснований, с которыми он образует комплекс в темноте (с неповрежденной ДНК фермент не связывается). На свету комплекс распадается, при этом происходит мономеризация димеров. В клетке эукариот фермент локализован в ядре, у прокариот — в непосредственной близости к нуклеоиду. В частности, он не обнаруживается в безнуклеоидных миниклетках, которые образуют некоторые мутанты Е. coli. Известен мутант phr E. coli, у которого блокирована фотореактивация. При облучении видимым светом у этого мутанта не исчезают тиминовые димеры из ДНК. Фермент фотореактивации найден у примитивных микроорганизмов, как микоплазмы, у многих высших растений и животных. Фермент образует стабильный комплекс с пиримидиновым димером и используя энергию поглощенног им света разрушает димер без разрыва цепей ДНК. Если под воздействием алкилирующих агентов - N-метил -N-нитрозомочевины или N1, N-диметилнитрозогуанидина, в ДНК образовался О6- метил- или О6-алкилзамещенные гуаниновые остатки, то деалкилирование таких остатков идет при участии ферментов - О6-метилгуанин-ДНК-алкилтрансферазы, которая катализирует перенос алкильных групп на сульфгидрильные группы цистеиновых остатков фермента, при этом акцепторный белок инактивируется. Это у бактерий и млекопитающих. Репарация однонитевых разрывов ДНК осуществляется последовательным действием 3'-фосфодиэстеразы, ДНК-полимеразы b и ДНК-лигазы [18]. Восстановление однонитевого разрыва ДНК происходит с использованием в качестве матрицы комплиментарной цепочки ДНК. Репарация данного типа повреждений может также происходить с участием механизма эксцизионной репарации нуклеотидов. Эксцизионная репарация осуществляется в несколько этапов. На первом этапе поврежденное основание удаляется ДНК гликозилазами, далее сахар без основания (АП-апуриновый-сайт) вырезается АП-эндонуклеазами. На втором этапе образовавшаяся брешь размером в одно основание восстанавливается до исходной последовательности с помощью совместного действия экзонуклеаз, полимеразы и ДНК лигазы
58. Эксцизионная репарация Эксцизионную репарацию, т. е. связанную с удалением поврежденного участка ДНК, называют также репарацией по типу выщепления — замещения или более образно «механизм режь — латай». Наиболее подробно изучена именно репарация ДНК, содержащей тиминовые димеры. Появление димеров приводит к локальной денатурации ДНК, что влечет за собой нарушение процесса репликации: каждый тиминовый димер в ДНК Е. coli задерживает репликацию на 10 с. Эксцизионная репарация представляет собой многоэтапный процесс и заключается в: 1) «узнавании» димера, 2) надрезании одной цепи ДНК вблизи димера — инцизии, 3) удалении димера — эксцизии, 4) ресинтезе ДНК и 5) восстановлении непрерывности репарируемой цепи за счет образования ковалентных связей сахарофосфатного скелета молекулы. «Узнавание» повреждения в ДНК осуществляет фермент УФ-эндонуклеаза, который реагирует не только на димеры тимина, но и на многие другие изменения, приводящие к локальному нарушению структуры ДНК. Эндонуклеаза ответственна и за инцизию, т. е. надрезание одной цепи ДНК (разрыв фосфодиэфирных связей) непосредственно около димера с 5'-конца в поврежденной цепи. Эксперименты in vitro с облученной ДНК показали, что число однонитевых разрывов оказывается равным числу димеров в молекуле.
Эксцизию, или вырезание димера из молекулы ДНК, осуществляет другая нуклеаза. Димер удаляется в составе короткого олигонуклеотида (3—5 оснований), что может сопровождаться дальнейшей деградацией поврежденной спирали. Продукты деградации облученной ДНК, содержащие тиминовые димеры, можно обнаружить в клетках. У некоторых бактерий димеры находили и в культуральной среде. Деградацию ДНК осуществляет АТФ-зависимая ДНКаза. В результате эксцизии и последующей деградации ДНК образуются однонитевые бреши, или пробелы. Ресинтез ДНК, в результате которого заполняются бреши, идет с использованием в качестве матрицы интактной цепи. Такой репаративный синтез ДНК напоминает «дополнительную» репликацию, обнаруженную в пахитене у эукариот. Основной фермент, ответственный за эксцизию димеров и репаративный синтез ДНК, — это ДНК-полимераза I, кодируемая геном pоl А. Тем не менее у мутантов pol А, дефектных по ДНК-полимеразе I, все же наблюдается остаточный репаративный синтез, который связан с активностью ДНК-полимеразы II. Известно, что неполуконсервативный синтез ДНК в 99% случаев происходит на коротких участках длиной до 30 нуклеотидов. За эту реакцию ответственна ДНК-полимераза I. В 1% случаев синтез идет на гораздо более длинных отрезках — 1000—1500 нуклеотидов
По-видимому, эту реакцию и осуществляет ДНК-полимераза II. Последний этап эксцизионной репарации заключается в восстановлении непрерывности репарируемой цепи ДНК с помощью фермента ДНК-лигазы, кодируемого геном lig E. coli. Температурочувствительные мутанты по этому гену не способны не только завершать процесс эксцизионной репарации в непермиссивных условиях, но и накапливают фрагменты Оказаки при репликации ДНК. 59. Неспаренные основания в ДНК могут возникать в результате трех событий: 1) прямого повреждения оснований ДНК или их предшественников продуктами клеточного метаболизма; 2) ошибочной подстановки некомплементарного основания ДНК-полимеразой в ходе репликации 3) рекомбинационной интеграции однонитевого участка ДНК в неабсолютно идентичную ДНК, партнера по рекомбинации. Все события приведут к образованию гетеродуплексной ДНК, которая и становится субстратом для ферментов, корректирующих неправильные пары оснований. Процесс репарации ДНК при обнаружении ферментами неканонической пары в дуплексе ДНК проходит через ряд реакций. Обычно такая цепь реакций начинается с обнаружения и удаления одного из нуклеиновых оснований неканонической пары в ДНК, катализируемого соответствующим ферментом группы AP-эндонуклеаз типа II (апуриновых/апиримидиновых эндонуклеаз) Затем происходит гидролитическое расщепление 3'-фосфоэфирной связи, причем фосфат остается в 5'-положении уходящей цепи ДНК. Далее с 3'-концом в месте расщепления цепи связывается ДНК-полимераза бета, приводящая к образованию шиффа между ферментом и 3'-цепью ДНК, катализирующая гидролитическое удаление дезоксирибозилфосфатного остатка (реакция В), таким образом освобождая 3'-конец для включения соответствующего канонической паре нуклеотида.
60.SOS-репарация относится к пострепликативной репарации. Этот тип репарации был открыт в клетках мутантов E. coli, не способных выщеплять тиминовые димеры. В таких клетках после ультрафиолетового облучения происходит репликация ДНК, хотя и медленнее, чем в клетках дикого типа. Показано, что в клетках мутантов uvr А после действия ультрафиолетового света синтезируется ДНК с однонитевыми пробелами, или брешами, причем длина вновь синтезированных фрагментов соответствует среднему расстоянию между возникшими в родительской ДНК тиминовыми димерами. Таким образом, после репликации нерепарированной ДНК против тиминовых димеров образуются бреши, которые, как оказалось, исчезают при последующей инкубации клеток в питательной среде. Этот тип репарации не происходит в клетках гес-мутантов, дефектных по рекомбинации. Поэтому постреплика-тивную репарацию называют также рекомбинационной репарацией. Механизм пострепликативной репарации наименее специфичен,так как здесь отсутствует этап узнавания повреждения. Представления об этом типе репарации связаны со знанием механизмарекомбинации. Рекомбинационная пострепликативнаярепарация — это быстрый способ восстановления нативнойструктуры по крайней мере части дочерних молекул ДНК. При этом тиминовые димеры остаются в исходных родительских нитях. Эта репарация происходит уже в первые минуты после облучения. Существует и другая разновидность — медленная пострепликативная репарация, для осуществления которой требуется несколько часов. Ее проводит система ферментов, которых нет в необлученных клетках и которую индуцирует облучение. Этот механизм получил наименование SOS-репарации. Его характерная черта — неточность восстановления первичной структуры ДНК, в связи с чем он получил также название репарации, склонной к ошибкам. При этом, по мнению ряда авторов, возможен репаративный синтез ДНК «в обход» тиминовых димеров, или, точнее, за счет использования в качестве матрицы цепи ДНК, содержащей димеры. Пострепликативная репарация существует не только у бактерий, но и в клетках эукариот. Она обнаружена и у млекопитающих, для которых получены данные о том, что пострепликативные бреши могут заполняться не за счет рекомбинации, а за счет синтеза ДНК de novo. Один из типов пигментной ксеродермы у человека (XPvar) связан с блоком пострепликативной репарации.
Дата добавления: 2015-12-16 | Просмотры: 907 | Нарушение авторских прав |