|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Мобильные генетические элементыС момента возникновения хромосомной теории наследственности до конца 70-х годов представление о том, что каждый ген имеет определенное место на хромосоме и не способен произвольно менять его, казалось незыблемым. Единственным известным способом перемещения генов друг относительно друга были хромосомные мутации — транслокации и инверсии. Другое очень распространенное и обоснованное представление гласит о том, что в геноме данного вида организмов содержится вполне определенное количество копий какого-либо конкретного гена. Изменение числа копий может также происходить в результате хромосомных мутаций — дупликаций и делений..В 40—50-х годах XX в. американская исследовательница Б. Мак-Клинток генетическими методами показала, что в хромосомах кукурузы предположительно существуют генетические элементы, способные перемещаться в геноме — исчезать с прежних мест и появляться в новых. Спустя четверть века американские и советские генетики независимо методами молекулярной биологии и генной инженерии доказали существование генетических элементов, способных к перемещению. Свойства мобильных генетических элементов. Мобильные генетические элементы обнаружены у самых разных организмов: у бактерий, дрожжей, растений и животных. Мобильные генетические элементы, выделяемые у разных видов, существенно отличаются по длине нуклеотидной последовательности и, следовательно, свойствам. У дрозофилы, хорошо изученной в этом отношении, описано около десяти типов мобильных элементов, одни из которых похожи друг на друга, иные резко отличаются. Количество копий, содержащихся в одном геноме, колеблется для разных мобильных элементов от К) до 1000. Длина нуклеотидной последовательности мобильных элементов варьирует в широких пределах, от 1 тыс. до 10 тыс. пар нуклеотидов. Наличие на концах длинных концевых повторов — типичная черта строения мобильных элементов. Способы перемещения. Существует, вероятно, не менее двух способов перемещения мобильных генетических элементов. Первый из них связан с «вырезанием» мобильного элемента в одном месте хромосом и встройкой его в другом месте. Такие перемещения, по-видимому, имеют обычно случайный, ненаправленный характер. Другой тип событий представляет собой направленные перемещения генетических элементов. В этом случае в исходном положении мобильный элемент сохраняется, но появляется и в новом, хотя и вполне определенном месте. Таким образом, в данном случае речь идет о появлении дополнительной копии мобильного элемента. Для этого необходимо удвоение молекулы ДНК данного мобильного элемента и его последующее встраивание в определенное место генома. Роль мобильных генетических элементов еще предстоит тщательно изучить. Однако в отдельных случаях уже имеется ясность. Так, Б. Мак-Клинток показала, что встраивание мобильного элемента рядом с геном, контролирующим окраску семян у кукурузы может включать этот ген. В результате окраска зерен изменяется, например с красной на белую. Прозорливость Б. Мак-Клинток состоит в том, что своими генетическими исследованиями она предсказала существование мобильных элементов в каком-либо гене или в непосредственной близости от него, что может приводить к резким наследуемым изменениям его состояния, т. е. по существу к появлению мутации. В последнее время стало ясно, что многие давно известные и хорошо изученные точечные мутации у дрозофилы, мыши и других организмов в действительности представляют собой результат встраивания или вырезания мобильных генетических элементов. 45% генома – подвижные (мобильные, или мигрирующие) элементы (ПЭ). Биологическая роль ПЭ. Проявляются как в онто-, так и в филогенезе. 1. Горизонтальный перенос устойчивости к антибиотикам, лекарствам, ядам – у бактерий. 2. Мутации генов за счет включения ПЭ в кодирующую часть генов. 3. Вмешательство в работу клеточных генов – изменение их активности (рак). 4. Перестройки хромосом, перенос генов и целых наборов генов в пределах одного генома и из одного генома (напр., вируса) в другой (хозяина). 5. Стабилилизация концов хромосом у дрозофил.
62. Общая схема гомолог. рекомбинации. Образ-е делеций и дупликаций в результ внутримолек и межмолек эктопической рекомбинации. Чаще всего рекомбинацию в узком смысле слова связ с кроссинговером,т.е. с перекомбинацией генов, локализ. в гомологичных хромосомах(т.е.гомологичная рекомбинация = кроссинговер). Разработана гипотеза «разрыв-воссоединение». Модель Холлидэя:Проц рекомбинации инициируют разрывы в нитях ДНК одинаковой полярности. Разрывы могут быть и не строго гомологичны. 1этап: молекулы ДНК, вступающие в рекомбинацию, образ.гибридные участки –так наз гетеродуплексы, в кот одна цепь происх от одной молек, а др- от другой. Это полухиазма.Затем она может видоизменяться путем миграции вдоль конъюгирующих хроматид(мол-л ДНК). Этот проц наз миграция ветвей полухиазмы. 2этап: в точке перекреста нити разрываются. При этом рвутся либо нити, в кот были первичные разрывы, либо другие две. Предполаг, что оба типа разрывов равновероятны. Т.о., получаются либо две нерекомбинантные по концевым маркерам молекулы(А-С, а-с), несущие гибридный участок-зону гетеродуплекса в районе средн маркера В\b, либо 2 мол,рекомбинантные по концевым маркерам и также гетеродуплексные в районе средн маркера. Поскольку, согласно положению Уотсона и Крика,мутации- это изменения чередования нуклеотидов в ДНК, аллели одного гена, в частности B/b, различаются по составу нуклеотидов(как мин по 1 паре оснований). Тогда в участке гетеродуплекса должна образоваться зона локального неспаривания оснований. Эти участки узнают спец ферменты репарации,обеспечивающие структ стабильность ДНК. Они устраняют неспаренное основание и заменяют его на комплементарное. В таком проц коррекции с равной вероятностью матрицей служит 1 из 2 нитей гетеродуплекса. Эктопическая рекомб. –кроссинговер между отдельно повторяющимися гомологичными последовательностями,разбросанными по геному. Внутрихромосомные перестройки(=абберации) подразделяют на дефишенси, или концевые нехватки; делеции — выпадения частей хромосомы, не захватывающие теломеру; дупликации, или удвоения (умножения) части хромосомы; инверсии — изменения чередования генов в хромосоме вследствие поворота участка хромосомы на 180°.Межхромосомные перестройки включают транслокации — перемещения части одной хромосомы на другую, не гомологичную ей. Делеции.Вследствие нехваток хромосомы укорачиваются, и физическое отсутствие участка одного из гомологов приводит к гемизиготному состоянию генов, находящихся в нормальном гомологе. Если теряются доминантные аллели одного из гомологов гетерозиготы, то наблюдается фенотипическое проявление рецессивных аллелей хромосомы, не затронутой аберрацией. Поскольку вследствие делеций теряются участки хромосом, у гетерозигот по этим..перестройкам наблюдаются характерные нарушения.Бoлee длинная хромосома образует петлю на участке, соответствующем делеции. Делеции обычно летальны в гомозиготе,что указывает на выпадение жизненно важных генов. Дупликации могут происх в пределах одной хр или сопровождаться переносом копии участка на др хр. Повторы, возникшие в одной хромосоме, могут располагаться тандемно (ABCBCDE...) или инвертировано(АВССВDE). По-видимому, главной причиной множественных повторов участков генетического материала является так наз неравный кроссинговер. Дупликации и делеции часто возникают в результате разрывов хромосом. Дупликации играют существенную роль в эволюции генома, поскольку они создают дополнительные участки генетического материала, функция которых может быть изменена в результате мутаций и последующего естественного отбора. 63. Сайт-специфическая рекомбинация. Геном фага γ проникает в бактериальную клетку в линейной форме, однако на концах линейной молекулы ДНК есть так называемые липкие концы — однонитевые участки по 12 нуклеотидов, комплементарные друг другу. В клетке ДНК γ замыкается в кольцо. В таком виде она интегрирует в геном бактерии. Кольцо генома А, реципрокно рекомбинирует с кольцом бактериальной хромосомы. Один обмен приводит к интеграции ДНК фага γ с ДНК бактериальной хромосомы. Интеграция профага может происходить только в одном месте на хромосоме Е. coli, названном att X (attachment site/.)- Аналогичный участок есть в геноме бактериофага. При этом рекомбинация происходит в отсутствие протяженной гомологии. Общим у фага и бактерии оказался участок всего в 15 п. н.: ...GCTTTTTTATACTAA... (показана только одна цепь ДНК) Вырезание (эксцизия) профага из хромосомы осуществляется по тому же механизму реципрокной, сайт-специфической рекомбинации. Как интеграцию, так и эксцизию профага Л контролируют два фаговых гена: int и xis.Сайт-специфическая рекомбинация происходит точно, но не безошибочно. Приблизительно один раз на 1 млн. при эксцизии профага рекомбинация осуществляется не в сайте, а захватывает участки gal или bio. Так возникают трансдуцирующие частицы, у которых часть генетического материала профага замещена генами бактерии. Во всех этих случаях в рекомбинацию вовлекаются те же последовательности из 15 пар нуклеотидов, которые встречаются в генах gal и bio. За пределами этих 15 п. н. гомология отсутствует. Сайт-специфическую рекомбинацию проводит фермент интеграза, кодируемый локусом int фага γ.
64. Транспозиция. Структурная и функциональная организация мобильных генетических элементов про- и эукариот. Классификация мутаций: внутрихромосомные, межхромосомные перестройки и промежуточное положение – транспозиции, т.к. они могут происходить как между негомологичными хромосомами, так и в пределах одной хромосомы. Транспозиция – перемещение небольших участков генетического материала в пределах одной хромосомы или между разными хромосомами. Происходят при участии особых подвижных или мигрирующих генетических элементов. Изучение молекулярной структуры мобильных генетических эл-тов на мутантной по 3 lac-генам Е.coli –1960. Общими для этих мутантов были вставки большей или меньей длины. Эти вставляемые в разные участки генома молекулы ДНК – IS-элементы (insertion sequenses). Особенности этих элементов: 1)на концах инвертированные повторы нуклеотидной последовательности (десятки пар нуклеотидов) 2) большинство Isэлементов содержит ген для фермента транспозазы, ответственных за их перемещение 3) могут содержать несколько сигналов начала и конца транскрипции 4) в точке внедрения каждого элемента всегда обнаруживается дупликация (4-9 пар нуклеотидов). Есть 3 мех-ма транспозиции для эукариот: 1) эксцизия предсуществующего транспозона с переносом на новое место – нерепликативная транспозиция. 2) репликация ДНК транспозона с последующей траспозицией - репликативная транспозиция 3) обратная транскрипция РНК-копии транспозона и перемещение ДНК-копии на новое место – РНК-опосредовання транспозиция. 65. Перемещаясь случайным образом, мобильные генетические элементы существенно влияют на структуру генетического материала хозяина и имеют фундаментальное значение в формировании генетической изменчивости. Считают, что транспозиционная активность МГЭ вызывает до 80 % спонтанных мутаций и является основной причиной их возникновения. Мобильные элементы оказывают различные регуляторные эффекты. ммуноглобулинов. 66. IS-элементы. В большинстве своем мобильные элементы прокариот и эукариот построены по сходному плану. Сами элементы состоят из центральной части, фланкированной инвертированными повторами (ИП). Центральная часть обычно содержит ген (или гены), кодирующие белки транспозиции. Главный белок транспозиции – транспозаза. Некоторые бактериальные транспозоны имеют на концах длинные ИП, в свою очередь являющиеся мобильными IS-элементами. В этих случаях центральная часть транспозона содержит только посторонние гены, а гены транспозиции находятся в IS-элементах, причем один из них инактивирован одной или более мутациями. Структура мобильных элементов определяет механизмы их перемещений. Хотя эти механизмы различаются в деталях, имеется общий принцип реакций транспозиции. Процесс происходит в 3 этапа. На первом этапе 2 молекулы транспозазы соединяются с концами подвижного элемента, сводят концы вместе и генерирует в них разрывы, чаще всего в обеих цепях. Затем транспозаза делает в обеих цепях ДНК-мишени ступенчатые разрывы, отстоящие друг от друга на столько пар нуклеотидов, сколько обнаруживается в ДПП данного элемента. Второй этап – обмен цепями, приводящий к рекомбинации между ДНК оставляя, за счет ступенчатости разрывов, бреши между 5'-P-концами элемента и 3'-OH-концами мишени. Катализируемое транспозазой расщепление и замыкание концов цепей ДНК происходит без потери энергии связей и не требует АТФ, что напоминает консервативную сайт-специфическую рекомбинацию. Отличие от последней заключается в том, что транспозаза не образует ковалентной связи с 5’-P концом ДНК. На третьем этапе происходит репаративный синтез брешей, формирующий ДПП, а иногда еще и репликация элемента. Таков общий общий механизм транспозиционной рекомбинации. IS-элементы: небольшие (размером не более 2,5 т.п.н.) элементы, которые состоят из центральной части с геном транспозазы, фланкированной двумя инвертированными повторами. Основные механизмы транспозиций(на рисунках):Репликативная транспозиция отличается тем, что мобильный элемент, перемещаясь в другую молекулу, оставляет свою копию в исходной ДНК. Это может произойти только за счет удвоения (репликации) элемента. При репликативной транспозиции на концах подвижного элемента происходят разрывы с образованием выступающих 3’-OH-концов. Одновременно транспозаза делает разрывы в ДНК-мишени. 3’-OH-концы подвижного элемента ковалентно связываются с 5’-Р-концами мишени, и образуется структура с двумя вилками репликации на концах подвижного элемента. В вилках репликации инициируется синтез ДНК (направленный «внутрь»). В результате образуется две копии мобильного элемента. При этом репликоны, содержащие «старую» и «новую» копию мобильного элемента сливаются (образуется коинтеграт). Коинтеграты разрешаются (разрезаются) на 2 репликона в рекомбинационном res -сайте ферментом резолвазой. Старая и новая копии мобильного элемента в коинтеграте находятся в одной ориентации, и разрешение коинтеграта идет через сложную фигуру, напоминающую восьмерку. В результате снова образуется 2 репликона, но теперь каждый из них несет копию мобильного элемента. Реакция относится к сайт-специфической рекомбинации. Репликативный механизм транспозиции распространен сравнительно мало. Он обнаружен у мобильного элемента Is6, фага Mu и бактериальных транспозонов семейства Tn3 с короткими ИП. Нерепликативная транспозиция заключается в вырезании элемента и его перемещении в новое место. При этом 2 молекулы транспозазы связываются с концами мобильного элемента и делают разрывы одновременно в обеих цепях ДНК на концах мобильного элемента и в ДНК-мишени. Далее транспозаза сводит вместе концы мобильного элемента и ДНК-мишень, 3-OH-концы элемента соединяются с 5-Р-концами ДНК-мишени, а между 3’-OH-концами ДНК-мишени и 5’-Р- концами элемента образуется брешь, которая заполняется с помощью репаративного синтеза ДНК, в результате чего на концах мобильного элемента возникают ДПП строго фиксированной длины. В исходном репликоне остается ДНР. Будет ли он репарирован – зависит хозяйской клетки. Этот механизм характерен для большинства мобильных элементов бактерий и эукариотических элементов с короткими ИП. По такому типу перемещаются многие IS-элементы и мобильные элементы, которые называют составными: Tn5, Tn9, Tn10 и другие. Составные транспозоны отличаются тем, что у них инвертированные повторы представлены IS-элементами, которые находятся в обратной или (гораздо реже, например, Tn9) в прямой ориентации.
Вопрос № 67 (дуплицированными прямыми повторами (ДПП), инвертированные повторы (ИП)) Собственно транспозоны несут кроме транспозазы другие гены, не имеющие отношения к транспозиции (чаще всего гены устойчивости к антибиотикам). Собственно транспозоны можно в свою очередь разделить на следующие группы 1) Сложные транспозоны (семейство Tn3) – короткие ИП на концах, делают в ДНК-мишени ДПП из 5 п.н. и перемещаются по механизму репликативной транспозиции. 2 ) Составные транспозоны (Tn5, Tn9, Tn10) с длинными ИП, представляющими собой различные IS-элементы. Длина ДПП обычно 9 п.н.
У самого Tn3 центральная часть содержит гены транспозазы, резолвазы и бета-лактамазы bla (обеспечивает устойчивость к антибиотикам пенициллинового ряда). Ген транспозазы tnA кодирует большой белок из примерно 1000 а.о., ген резолвазы tnR кодирует белок из 185 а.о. Гены транспозазы и резолвазы транскрибируются в противоположных направлениях с промоторов, расположенных в межгенном пространстве длиной 170 п.н. В межгенном пространстве находится и сайт res, по которому происходит разрешение коинтегратов. Транскрипции генов резолвазы и транспозазы конкурируют друг с другом, и ген резолвазы выступает как ген-регулятор гена транспозазы. К семейству Tn3 относятся Tn1, Tn1000 и др. Нерепликативная транспозиция заключается в вырезании элемента и его перемещении в новое место. При этом 2 молекулы транспозазы связываются с концами мобильного элемента и делают разрывы одновременно в обеих цепях ДНК на концах мобильного элемента и в ДНК-мишени. Далее транспозаза сводит вместе концы мобильного элемента и ДНК-мишень, 3-OH-концы элемента соединяются с 5-Р-концами ДНК-мишени, а между 3’-OH-концами ДНК-мишени и 5’-Р- концами элемента образуется брешь, которая заполняется с помощью репаративного синтеза ДНК, в результате чего на концах мобильного элемента возникают ДПП строго фиксированной длины. В исходном репликоне остается ДНР. Будет ли он репарирован – зависит хозяйской клетки. Этот механизм характерен для большинства мобильных элементов бактерий и эукариотических элементов с короткими ИП. По такому типу перемещаются многие IS-элементы и мобильные элементы, которые называют составными: Tn5, Tn9, Tn10 и другие. Составные транспозоны отличаются тем, что у них инвертированные повторы представлены IS-элементами, которые находятся в обратной или (гораздо реже, например, Tn9) в прямой ориентации.
Вопрос №69
1) Ретротранспозоны с длинными прямыми концевыми повторами (ДКП) (класс I.1). Их структура соответствует ДНК-копиям геномов ретровирусов позвоночных, которые также являются мобильными элементами. 2) Ретроэлементы (класс I.2), не содержащие повторов на концах (некоторые авторы используют для них название «ретропозоны»). Ретровирусы являются «прототипами» ретротранспозонов. У ретроэлементов с ДКП транспозиция происходит по схеме, включающей РНК-интермедиат. С геномной ДНК элемента транскрибируется РНК-копия, но уже с короткими концевыми повторами, с нее путем обратной транскрипции синтезируется ДНК-копия с ДКП, которая встраивается в новое место с помощью интегразы.
Интегразы ретротранспозонов, несмотря на различие в названиях, полностью соответствуют транспозазам. Рекомбинация у ретроэлементов без концевых повторов менее изучена, но она также осуществляется через РНК-интермедиат. Другая группа ретротранспозонов – элементы класса I.2 (ретропозоны). Их размер – тоже около 5-6 т.п.н., но на концах они не имеют повторов. На 3’-конце они содержат небольшую последовательность поли-A. Прямых повторов в ДНК-мишени они либо не образуют, либо делают не всегда, и, если делают, то нерегулярной длины. Ретротранспозоны класса II можно разделяют на 2 типа: LINE (long interspersed nuclear elements) и SINE (short interspersed nuclear elements) – длиной 200-300 п.н., которые не кодируют никаких белков и не способны к самостоятельному перемещению, а перемещаются, по-видимому, за счет элементов LINE. Механизм перемещения LINE- и SINE-элементов представлен на рисунке. В отличие от ретротранспозонов I типа, здесь реакцию интеграции в хозяйский геном инициируетет РНК-копия элемента. Эндонуклеаза делает ступенчатые ОНР в ДНК-мишени и РНК-копия прикрепляется к концу ДНК-мишени в точке разрыва. На матрице РНК-копии с помощью обратной транскриптазы строится ее ДНК-копия. Свободная группа 3’-OH в точке разрыва используется как праймер для обратной транскриптазы. Потом РНК-копия удаляется с помощью РНКазы H, клеточная репаративная система достраивает вторую цепь ДНК, которая оказывается интегрирированной в реципиентную ДНК. При этом на концах встроенного элемента могут возникать ДПП различной длины. Дата добавления: 2015-12-16 | Просмотры: 1192 | Нарушение авторских прав |

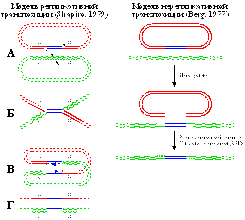 Транспозон Tn3 представляет семейство мобильных элементов с короткими ИП (35-50 п.н.), перемещающимися с помощью репликативной транспозиции и образующими ДПП из 5 п.н.
Транспозон Tn3 представляет семейство мобильных элементов с короткими ИП (35-50 п.н.), перемещающимися с помощью репликативной транспозиции и образующими ДПП из 5 п.н.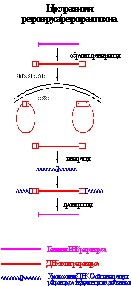 У эукариот широко распространены ретротранспозоны, в транспозициях которых задействованы фермент обратная транскриптаза (ревертаза) и РНК-копия элемента в качестве интермедиата. Ретроэлементы подразделяются на 2 группы:
У эукариот широко распространены ретротранспозоны, в транспозициях которых задействованы фермент обратная транскриптаза (ревертаза) и РНК-копия элемента в качестве интермедиата. Ретроэлементы подразделяются на 2 группы: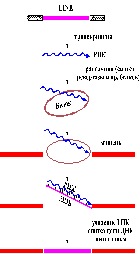 Интеграция ретротранспозонов с ДКП происходит по механизму, идентичному с нерепликативной транспозицией у прокариот.
Интеграция ретротранспозонов с ДКП происходит по механизму, идентичному с нерепликативной транспозицией у прокариот.