|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Фотосистемы I и II (ФС)Впервые идею о существовании двух фотосистем высказал американский биохимик Роберт Эмерсон в 1959 г. Он экспериментировал с хлореллой по изучению скорости фотосинтеза, используя показатель квантового выхода фотосинтеза:
Данные этого эксперимента легли в основу гипотезы о существовании двух фотореакций, которые индуцируются красным светом различной длины волны. Эта гипотеза в 60-х гг была подтверждена экспериментально. Из тилакоидов мембран были выделены компоненты ФСI и ФСII. Каждая из этих фотосистем имеет собственные ССК, реакционный центр и свой отрезок ЭТЦ. Компоненты ЭТЦ организованы последовательно в тилакоидных мембранах. Все переносчики содержат функциональные группировки, способные к окислительно-восстановительным превращениям. Одни переносчики электронной природы, другие – атомной природы. Различаются переносчики степенью сродства к электронам, о которой судят по окислительно-восстановительному потенциалу (Е) – это количественная мера сродства к электронам, измеряется в вольтах и относится к паре. Разные окислительно-восстановительные пары обладают разной степенью сродства к электронам, величина окислительно-восстановительного потенциала изменяется от – до +, место расположения переносчиков в ЭТЦ определяется величиной окислительно-восстановительного потенциала. Характеристика основных переносчиков 1) Цитохромы – электронные переносчики, функциональной группировкой которых является железо. Цитохромы представляют собой гемопротеины. Существуют разные цитохромы: а, b, c, d; они отличаются друг от друга по структуре гема и по аминокислотному составу. В ЭТЦ фотосинтеза есть цитохром b и цитохром c (f). 2)
В ЭТЦ фотосинтеза три вида хинонов: мембрансвязанные хиноны QA и QB, липидорастворимый, свободно мигрирующий по тилакоидной мембране пластохинон (Пх). 3) Пиридиннуклеотиды (НАД, НАДФ) – атомные переносчики. Окислительные цепи используют НАД, а восстановительные – НАДФ. 4) Флавопротеиды (ФАД) – атомные переносчики. ФАД входит в состав оксидоредуктазы, которая восстанавливает НАДФ. 5) Железопротеиды – электронные переносчики, функциональной группировкой которых является железо в негеминовой (в отличие от цитохромов) форме. К ним относят железо-серные белки (Fe-S) и ферредоксин (Фд). 6) Медьпротеиды – электронные переносчики. В ЭТЦ фотосинтеза представлены пластоцианином (Пц). 7) Марганецпротеиды – электронные переносчики. Входят в состав водорасщепляющего комплекса. Схема ЭТЦ предложена учеными Хиллом и Бенданом в 1960 г., она получила название Z-схема. Фотосистема I. В реакционном центре фотосистемы I находится димерная форма хлорофилла а с максимумом поглощения 700 нм (Р700). Первичным акцептором электронов (А1) является специфическая мономерная форма хлорофилла а. В роли вторичного акцептора электронов выступает железо-серный белок (А2 = Fe-S). Со вторичного акцептора электроны последовательно поступают на две формы ферредоксина (Фд): мембрансвязанную и водорастворимую. Ферредоксин образует комплекс с соединением, простетической группой которого является ФАД. ФАД-содержащая оксидоредуктаза, в свою очередь, способна восстанавливать НАДФ+. Фотосистема II. В реакционном центре – димерная форма хлорофилла а с максимумом поглощения 680 нм (Р680). Первичный акцептор – безмагниевый аналог хлорофилла феофитин (Фф). Восстановленный феофитин передает электроны на пластохинон. Пластохинон находится в тилакоидах в количестве, превышающем концентрации всех других переносчиков, и образует пул пластохинонов (QA, QB, Пх). Восстановление пластохинонов происходит после присоединения двух электронов. В восстановленном виде пластохиноны проявляют очень высокое сродство к протонам, которые поступают из стромы хлоропласта. Пластохиноны обеспечивают связь фотосистем I и II через цитохромный комплекс, который включает цитохром b6, белок Риске (Fe-S) и цитохром c (f). От цитохромного комплекса электроны идут на пластоцианин (Пц). Помимо прочих в фотосистему II входят следующие кофакторы: Са2+, Сl-, СО22-. Особняком стоит участок ЭТЦ, который передает электроны на восстановление окисленного Р680 от воды, так называемый, водорасщепляющий комплекс. При передаче электронов с Р680 на феофитин образуется окисленный хлорофилл, который обладает исключительно высоким сродством к электронам и является самым сильным биологическим окислителем. Благодаря этому происходит фотолиз воды, его осуществляет водорасщепляющий комплекс, который в активном центре содержит группу ионов марганца. Ионы марганца служат донором электронов для восстановления окисленного Р680. Отдавая электроны, ионы марганца становятся накопителями положительных зарядов, которые участвуют в окислении воды. После четырехкратного срабатывания в водорасщепляющем комплексе накапливается 4 сильных окисленных эквивалента в виде двух окисленных ионов марганца. Эти ионы взаимодействуют с двумя молекулами воды, отнимая от них 4 электрона, тем самым, вызывая разложение воды. Фотосистема II является наиболее уязвимым звеном фотосинтетического аппарата, который чаще всего повреждается в стрессовых ситуациях. Молекулярные механизмы ключевого этапа фотолиза воды до сих пор не ясны.
Известны следующие виды фосфорилирования: 1) окислительное; 2) субстратное (осуществляется посредством киназ); 3) фотофосфорилирование. Фотофосфорилирование – это процесс образования АТФ в ходе световой фазы фотосинтеза за счет энергии выделения электронов. Известны разные пути фотофосфорилирования: 1) Нециклический путь связан с нециклическим транспортом электронов от воды к НАДФ+: Н2О + НАДФ+ + АДФ + Фн → НАДФН2 + АТФ + О2↑ 2) Циклический путь. Иногда электроны от ферредоксина идут не на ФАД, а на восстановленный пластохинон. В этом случае НАДФ+ не восстанавливается, но протонный градиент создается и синтез АТФ идет. Часть хлоропластных белков синтезируется непосредственно в хлоропластах, для этого нужна АТФ, поэтому даже при нормальной длине волны иногда происходит переключение на циклический путь фотофосфорилирования. 3) Псевдоциклический путь. Иногда оксидоредуктаза восстанавливает не НАДФ+, а кислород, в результате этого образуется вода.
В процессах, связанных с окислительно-восстановительными реакциями, происходит изменение свободной энергии, которое рассчитывается по формуле Нернста:
Δσ° = – nFΔE0,
где Δσ° – изменение свободной энергии; n – количество переносимых электронов (n = 2); F – число Фарадея (F = 23 ккал/моль);
ЕНАДФ /НАДФН = – 0,32 В (электроноакцепторная система)
Δσ° = – 2 · 23 (– 0,32 – 0,82) = 52,44 ккал В процессе передачи электронов с воды на НАДФ+ энергия поглощается. Для образования одной макроэргической связи АТФ требуется 7,3 ккал/моль. Таким образом, во время фотофизического и фотохимического этапов световой фазы фотосинтеза происходит запасание 52,44 ккал энергии, часть этой энергии расходуется на синтез АТФ в ходе энзиматического этапа. Дата добавления: 2015-12-15 | Просмотры: 2143 | Нарушение авторских прав |
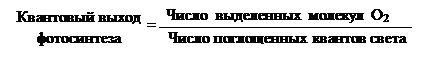
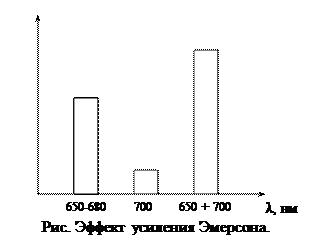 Используя красный свет разной длины волны Эмерсон обнаружил, что при освещении организма светом с длиной волны 650 – 680 нм выделяется определенное количество кислорода, при освещении водоросли красным светом с длиной волны 700 нм выделяется меньшее количество кислорода. При совместном освещении хлореллы длинно- и коротковолновым светом интенсивность фотосинтеза выше, чем при использовании света одной длины волны. Это повышение интенсивности фотосинтеза назвали эффектом усиления Эмерсона.
Используя красный свет разной длины волны Эмерсон обнаружил, что при освещении организма светом с длиной волны 650 – 680 нм выделяется определенное количество кислорода, при освещении водоросли красным светом с длиной волны 700 нм выделяется меньшее количество кислорода. При совместном освещении хлореллы длинно- и коротковолновым светом интенсивность фотосинтеза выше, чем при использовании света одной длины волны. Это повышение интенсивности фотосинтеза назвали эффектом усиления Эмерсона.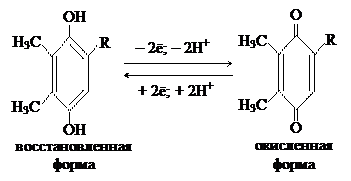 Хиноны – атомные переносчики, производные бензохинона:
Хиноны – атомные переносчики, производные бензохинона: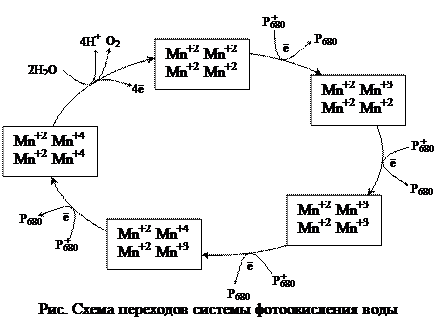
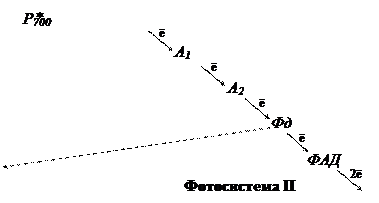



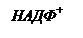

 ΔE0 – изменение окислительно-восстановительного потенциала (разность окислительно-восстановительного потенциала электронодонорной и электроноакцепторной систем).
ΔE0 – изменение окислительно-восстановительного потенциала (разность окислительно-восстановительного потенциала электронодонорной и электроноакцепторной систем).