|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
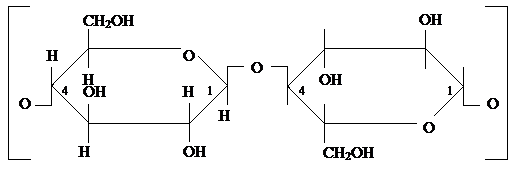
Клеточная стенкаНаличие клеточной стенки является исключительным признаком, который отличает растительные клетки от животных. Клеточные стенки растительных клеток имеют большое значение с общебиологической точки зрения, поскольку: 1) клеточная стенка определяет морфологию, размер и функции растительных клеток; 2) клеточная стенка участвует в регуляции растяжения, обеспечивая рост растения. Химическая и физическая структура клеточных стенок различается у разных клеток, но имеются два общих свойства: 3) Клеточные стенки могут состоять из различного числа слоев. 4) Вторичная клеточная стенка откладывается изнутри (между плазмалеммой и первичной клеточной стенкой), поскольку материал для ее построения поставляется из цитоплазмы. Химический состав клеточной стенки Все компоненты делятся на 4 группы: 1) Структурные компоненты, представленные у большинства автотрофных организмов целлюлозой (у некоторых водорослей этот маннаны и ксиланы). 2) Компоненты, образующие матрикс клеточной стенки представлены гемицеллюлозами, пектинами и белками. 3) Компоненты, инкрустирующие (пронизывающие) клеточную стенку представлены лигнином и суберином. 4) Компоненты, акрустирующие (покрывающие) клеточные стенки эпидермальных клеток представлены кутином и восками. Кроме того, клеточные стенки могут содержать большое количество минеральных веществ (например, силиката кальция и др.). 1) Структурные компоненты. На долю целлюлозы в первичной клеточной стенке приходится до 30 %, а во вторичной клеточной стенке – до 60 %. Это полимер β-глюкозы. Однако в настоящее время мономером целлюлозы считают целлобиозу, т. к. в первом каталитическом центре фермента, отвечающего за синтез целлюлозы, образуется целлобиоза, а в другом каталитическом центре она пришивается к растущей цепи молекулы целлюлозы.
Молекулы целлюлозы никогда не встречаются в виде одиночных цепей, а объединяются в мицеллы, которые, в свою очередь, объединяются в микрофибриллы, и далее – в макрофибриллы. За счет поочередного расположения кислородных мостиков в молекуле целлюлозы (то сверху, то снизу) создается наибольшее количество водородных связей между отдельными цепями. Молекулы целлюлозы в клеточной стенке располагаются очень упорядоченно. В центре макрофибриллы молекулы образуют кристаллическую решетку, эта часть носит название ядро (кор) макрофибриллы. Вокруг ядра молекулы располагаются хоть и параллельно, но не так упорядоченно, поскольку здесь между цепями целлюлозы внедряются молекулы гемицеллюлоз. Это место носит название кортекс (паракристаллическая часть). Гемицеллюлозы также сшивают отдельные макрофибриллы между собой. Особенности строения целлюлозы приводят к тому, что она очень стабильна к действию ферментов и химических реагентов. Гидролиз целлюлозы возможен лишь при длительном нагревании с концентрированной кислотой. У некоторых бактерий и растений есть фермент целлюлаза, которая способна разрушать 1-4 связи. Мономеры целлюлозы повторно не используются, поэтому деятельность целлюлаз проявляется лишь иногда (формирование отделительного слоя при листопаде). Целлюлозные фибриллы синтезируются белковыми комплексами, встроенными в цитоплазматическую мембрану. Эти комплексы, называемые розетками, являются элементами фермента целлюлозосинтетазы. Этот фермент имеет два каталитических центра: в первом происходит образование целлобиозы, а во втором эта целлобиоза пришивается к растущей цепи целлюлозы. Для каждого каталитического центра требуется наличие УДФ-глюкозы. Источником глюкозы является сахароза. Глюкоза переносится на УТФ, при этом третий остаток фосфорной кислоты отщепляется и вместо него пришивается глюкоза. В таком активированном состоянии глюкоза идет на синтез целлобиозы. 2) Матрикс. Фибриллы целлюлозы помещены в желеобразную массу – матрикс. Гемицеллюлозы – это производные пентоз и гексоз, небольшие по размерам (150 – 300 мономерных остатков). Мономеры соединяются между собой связями 1-4 и 1-6, т. о. гемицеллюлозы подобны разветвленной форме крахмала и гликогену. Пектиновые вещества являются преимущественно полимерами галактуроновой кислоты, которая представляет собой галактозу с группой – СООН у 6 углеродного атома. Остатки мономеров соединены связями 1-4. Отдельные цепи этих полимеров могут соединяться между собой с помощью ионов Са2+, который встают на место атомов водорода карбоксильных групп. Образованные соединения называются пектаты кальция. Если растение обитает в среде, где мало кальция, то может происходить мацерация клеток, поскольку именно пектаты кальция являются главным компонентом срединной пластинки. Ионы кальция, входящие в состав матрикса могут обмениваться на другие катионы, обеспечивая катионообменную способность клеточных стенок, что важно для клеток корневой системы. Белки матрикса. Наиболее часто встречается белок экстенсин, который по химической природе относится к гликопротеинам. Его аминокислотная часть на 20 % представлена пролином и гидроксипролином и содержит значительную часть серина и треонина. Углеводная часть представлена тетрасахаридом арабинозой. Роль этого белка заключается в том, что благодаря нему клеточные стенки могут в определенной степени быть эластичными. Содержание этого белка в клеточной стенке составляет около 10 %. 3) Инкрустирующие компоненты. Основным инкрустирующим компонентом является гетерополимерное ароматическое соединение лигнин. Интенсивная лигнификация начинается после прекращения роста клетки. Отложение лигнина увеличивает механическую прочность клеточной стенки. Мономерами лигнина являются ароматические спирты. Наиболее часто встречаются кумаровый и синаповый спирты:
Полимеры лигнина образуются путем конденсации предшественника – шикимовой кислоты, которая является производным пентозофосфатного пути окисления глюкозы. Синтез полимера происходит в межфибриллярном пространстве с участием пероксидаз. За счет гидроксильных группировок мономеров лигнин связывается водородными связями с пектиновыми веществами матрикса, т. о. лигнин заполняет межфибриллярное пространство клеточной стенки. Лигнин является очень стойким соединением, он разрушается лишь ферментами, которые синтезируется некоторыми грибами. От наличия или отсутствия этих грибов зависит содержание гумуса в почве. Суберин – полимер моно-, ди- и триокси- жирных насыщенных и ненасыщенных кислот с количеством атомов углерода 16 – 18 (преимущественно ненасыщенных). По природе это гидрофобное вещество, поэтому пропитанные им клеточные стенки становятся трудно проницаемыми для воды и растворов. 4) Акрустирующие компоненты. Акрустирующие вещества по своей природе гидрофобны. Они покрывают поверхность эпидермальных клеток. Основное вещество – кутин, сходный по химическому составу с суберином, но мономерами его являются преимущественно насыщенные кислоты. Предшественник кутина секретируется на поверхность эпидермальных клеток, превращаясь в кутин в процессе окислительной полимеризации. Кроме кутина могут секретироваться воска – сложные эфиры жирных кислот и высокомолекулярных одноатомных спиртов. По природе гидрофобны. Дата добавления: 2015-12-15 | Просмотры: 818 | Нарушение авторских прав |