|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Электрофизиологическая система регуляцииЭлектрофизиологическая регуляция включает возникновение градиентов биоэлектропотенциалов между различными частями растения благодаря различию величин мембранного потенциала в клетках разных тканей, зон и органов растений. Эти градиенты не остаются постоянными, а совершают периодические колебания, обуславливающие изменения условий внутренней и внешней среды. Различают потенциал покоя и потенциал действия. Потенциал покоя – это мембранный потенциал, существующий у живой клетки в состоянии физиологического покоя, он возникает за счет разности концентрации анионов и катионов. В норме его величина варьирует в пределах 60 – 90 милливольт (максимальная величина, которую выдерживает мембрана составляет 200 мВ). Потенциал действия – быстрые колебания мембранного потенциала, возникающие при возбуждении некоторых растительных клеток. Этот сигнал обеспечивает достаточно быструю передачу информации о действии раздражителя в организме. Потенциал действия подчиняется правилу «Все или ничего», т. е. возникает при достижении раздражителя некоторого порогового значения. Потенциал действия возникает через влияние на проницаемость мембран, увеличивая выход ионов хлора приводит к деполяризации мембран. Клетки при этом стремятся реполяризовать мембраны, что достигается не обратным ходом ионов Cl-, а выходом ионов К+. В результате мембранный потенциал восстанавливается, но уже с другим распределением ионов. Потенциал действия, возникнув на определенном участке мембраны, распространяется от клетки к клетке по плазмодесмам со скоростью 0,08 – 0,5 м/сек. Все межклеточные системы регуляции взаимосвязаны. Фитогормоны могут активировать или ингибировать транспорт трофических факторов. Межклеточные системы регуляции девствуют через внутриклеточные. На уровне организма при сохранении его целостности сохраняется взаимосвязь всех регуляторных механизмов. Эти системы регуляции включены как у устойчивых, так и у неустойчивых растений. Неустойчивые растения характеризуются тем, что переключение метаболизма происходит очень быстро, это затрудняет регуляторные объединения защитных механизмов, вследствие чего снижается сбалансированность процессов. У устойчивых растений в процессе эволюции сформировалась способность к постепенному и последовательному взаимодействию всех систем регуляции, что и обеспечивает длительность существования организмов в неблагоприятных экологических условиях. ТЕМА 3. Фотосинтез Фотосинтез – это процесс биологического преобразования лучистой энергии Солнца в энергию химических связей органических соединений. Значение фотосинтеза: 1) Обеспечение энергией всего живого на планете. Ежегодно, благодаря процессу фотосинтеза, на планете образуется 200 млрд. тонн биомассы. Это количество эквивалентно 3 · 1021 Дж энергии. Человечество ежегодно потребляет около 1 млрд. тонн биомассы, что эквивалентно 15 · 1018 Дж и составляет около 0,5% энергии, запасенной в процессе фотосинтеза. Общее же потребление энергии в мировом масштабе составляет 3,5 · 1020 Дж, что составляет около 10% энергии, запасенной в процессе фотосинтеза. Разведанные запасы ископаемых природных источников соответствуют запасу энергии получаемой при фотосинтезе примерно за 100 лет. Таким образом, фотосинтез лежит в основе энергообеспечения всей биосферы. 2) В результате фотосинтеза как побочный продукт выделяется кислород. Это привело к революционным преобразованиям на планете: ü восстановленная среда заменилась на окисленную; ü на смену анаэробным формам жизни пришли аэробные; ü первичная разреженная атмосфера превратилась во вторичную плотную азотно-кислородную; ü в верхних слоях атмосферы образовался озоновый экран, который стал фильтровать солнечное излучение. В настоящее время фотосинтез лежит в основе биологического круговорота энергии и вещества, от которого зависит жизнь, ее разнообразие и возможности. 3.1. История изучения фотосинтеза До XVII в. существовала гипотеза почвенного питания растений. Это заключение основывалось на бытовых наблюдениях – чем темнее почва, тем она более плодородная. В XVII в. возникло представление о водном питании растений. В основу этого представления легли эксперименты Яна Батиста Ван-Гельмонта. Он взял веточку ивы, взвесил ее и посадил в большой глиняный горшок в почву, которую тоже предварительно взвесил. Через 5 лет он вновь повторил взвешивание. За это время масса ивового куста увеличилась на 74 кг, а масса почвы уменьшилась на 57 грамм. Ван-Гельмонт сделал вывод о том, что все необходимые элементы растения получают из воды. Водная теория была опровергнута во второй половине XVIII в., в 1772 году английским химиком Дж. Пристли был открыт фотосинтез. За 18 лет до этого Ломоносов высказал любопытные мысли о питании растений, он впервые заговорил о воздушном питании растений. Пристли в своих опытах смоделировал городскую атмосферу: в закрытом сосуде он сжигал свечу, затем он искал пути исправления воздуха, испорченного горением свечи. Под колпаком он оставил веточку мяты и заметил, что растение может исправить воздух, испорченный горением свечи. Результат своего эксперимента он донес до людей, и они стали активно разводить комнатные растения. Ян Ингенхауз провел около 500 экспериментов, на основе которых сделал вывод, что растения исправляют воздух, но только на свету и только зелеными частями. Он предположил, что в процессе фотосинтеза поглощается СО2. В 1782 году швейцарский пастор Жорж Сенебье подтвердил это предположение и назвал этот процесс углеродным питанием растений. Теодор Сюссор в 1804 году с помощью количественных методов показал, что растения поглощают СО2 и выделяют эквивалентное количество О2. Но нарастание сухой массы растений превышало количество поглощенного СО2, на основании этого был сделан вывод о том, что кроме углекислого газа поглощается еще и вода. В 1817 году Каванту и Пельтье случайно выделили хлорофилл. В 1865 г. в ходе эксперимента немецкий физиолог Сакс установил, что на свету в листьях образуется крахмал. Для этого он предварительно выдерживал растение в темноте, а затем выставлял его на свет. Через некоторое время он обесцвечивал лист спиртом и проводил качественную реакцию на крахмал. Общее уравнение фотосинтеза впервые предложил француз Бухтенгор:
СО2 + Н2О → [СН2О] + О2↑
СО2 + Н2S → [СН2О] + S↓
Он предложил общее уравнений фотосинтеза:
СО2 + Н2А → [СН2О] + А,
где Н2А – это какое-либо восстановленное соединение. Энергия света при этом тратится на разложение этого восстановленного соединения. Это предположение было подтверждено в 1937 г. Робертом Хиллом, который обнаружил, что изолированные хлоропласты на свету способны разлагать воду и выделять при этом кислород в присутствии акцепторов электрофонов и протонов. Он предложил следующую схему разложения воды:
4Н2О → 4ОН- + 4Н+ 4ОН- → 2Н2О + О2↑ + 4ē
В качестве акцептора протонов и электронов он использовал бензохинон:
Схему фотосинтеза можно представить следующим образом:
В целом, фотосинтез состоит из двух фаз: 1) Фаза преобразования энергии:
12Н2О → 24[Н+] + 6О2
2) Фаза превращения веществ:
6СО2 + 24[Н+] → С6Н12О6 + 6Н2О
Суммарное уравнение фотосинтеза:
6Н2О + 6СО2 → С6Н12О6 + 6Н2О
Существование двух фаз фотосинтеза было доказано в эксперименте с мигающим светом. В результате этого эксперимента было установлено, что больший выход продуктов при одинаковом количестве квантов возникал при импульсном освещении, а не при непрерывном. В 1954 г. американский биохимик Арно окончательно решил вопрос о существовании двух этапов фотосинтеза. Он установил, что выделенные хлоропласты под действие м света способны восстанавливать окисленный НАДФ+ и фосфорилировать АДФ. В 1958 г. он же установил, что гомогенат, лишенный мембран, полученный из стромы хлоропластов, способен ассимилировать углекислый газ, если в среде присутствуют НАДФ восстановленный и АТФ. Световая и темновая фазы фотосинтеза разделены во времени и пространстве. Световая фаза идет только на свету в третьей мембранной структуре хлоропластов (тилакоидах), темновая фаза протекает в строме хлоропластов и не зависит от света. Каждая фаза делится на этапы.
|
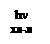
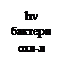 Термин «фотосинтез» предложен немцем Пфеффером в 1877 г. В то время общепринятым было мнение, что на свету происходит разложение углекислого газа до углерода, который присоединяется к воде, и кислорода, который выделяется в атмосферу. Иную трактовку процесса фотосинтеза выдвинул в 1893 г. русский биохимик Бах. Он высказал, что фотосинтез является сопряженным окислительно-восстановительным процессом, происходящим за счет энергии солнечного света, а кислород выделяется из воды через ряд промежуточных перекисных соединений. Но экспериментального подтверждения этому не было. В 30-х гг. ХХ в. вновь вернулись к вопросу о том, откуда выделяется кислород при фотосинтезе: из СО2 или из Н2О? Голландский микробиолог Карнелиус Ван-Ниль, будучи студентом III курса, занимался бактериальным фотосинтезом. Серные пурпурные бактерии способны осуществлять синтез органических веществ в присутствии сероводорода, но при этом не происходит выделение кислорода, а в клетках бактерий накапливаются частицы серы:
Термин «фотосинтез» предложен немцем Пфеффером в 1877 г. В то время общепринятым было мнение, что на свету происходит разложение углекислого газа до углерода, который присоединяется к воде, и кислорода, который выделяется в атмосферу. Иную трактовку процесса фотосинтеза выдвинул в 1893 г. русский биохимик Бах. Он высказал, что фотосинтез является сопряженным окислительно-восстановительным процессом, происходящим за счет энергии солнечного света, а кислород выделяется из воды через ряд промежуточных перекисных соединений. Но экспериментального подтверждения этому не было. В 30-х гг. ХХ в. вновь вернулись к вопросу о том, откуда выделяется кислород при фотосинтезе: из СО2 или из Н2О? Голландский микробиолог Карнелиус Ван-Ниль, будучи студентом III курса, занимался бактериальным фотосинтезом. Серные пурпурные бактерии способны осуществлять синтез органических веществ в присутствии сероводорода, но при этом не происходит выделение кислорода, а в клетках бактерий накапливаются частицы серы:

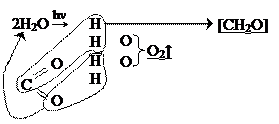
 Темновая фаза:
1) Карбоксилирование.
2)
Темновая фаза:
1) Карбоксилирование.
2) При использовании материала ссылка на сайт medlec.org обязательна! (0.092 сек.) |