|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
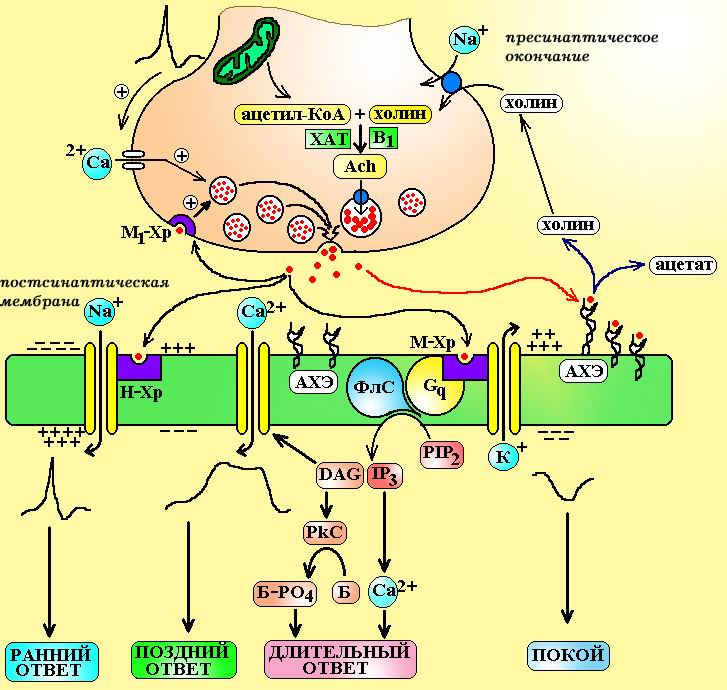
Основные этапы холинергической передачи и их фармакологическая коррекция1. Синтез и депонирование медиатора. Ацетилхолин синтезируется в пресинаптических окончаниях из ацетил-КоА и холина. В цитоплазме пресинаптического окончания содержится большое количество митохондрий, здесь путем окислительного декарбоксилирования a-кетокислот синтезируется ацетил-КоА. Холин поступает в клетку извне благодаря специальному трансмембранному переносчику. Транспорт холина в нейрон сопряжен с переносом ионов натрия и может быть блокирован гемихолином. Таблица 2. Сравнительная характеристика холинорецепторов клетки.
Примечание: a-бунгаротоксин – яд тайваньской гадюки Bungaris multicintus и кобры Naja naja. PTMA – фенилтриметиламмоний DMPP – диметилфенилпиперазин HHSDP – гексагидросиладифенол АВУ – атриовентрикулярный узел САУ – синоаурикулярный узел Синтез ацетилхолина проводит особый фермент холинацетилтрансфераза, путем ацетилирования холина. Образовавшийся ацетилхолин поступает в везикулы при помощи антипортера переносчика в обмен на протон. Работа этого переносчика может быть заблокирована векзамиколом. Обычно в каждой везикуле содержится от 1.000 до 50.000 молеукл ацетилхолина, а общее число везикул в пресинаптическом окончании достигает 300.000. 2. Выделение медиатора. Во время фазы покоя, через пресинаптическую мембрану выделяются единичные кванты медиатора (изливается содержимое 1 везикулы). Одна молекула ацетилхолина способна вызвать изменение потенциала мембраны всего на 0,0003 мВ, а то количество, которое содержится в 1 везикуле – на 0,3-3,0 мВ. Такие миниатюрные сдвиги не вызывают развития биологического ответа, но поддерживают физиологическую реактивность, тонус ткани-мишени. Активация синапса происходит в тот момент, когда на пресинаптическую мембрану приходит потенциал действия. Под влиянием потенциала мембрана деполяризуется и это вызывает открытие воротного механизма медленных кальциевых каналов. По этим каналам ионы Са2+ поступают в пресинаптическое окончание и взаимодействуют с особым белком в мембране везикул – синаптобревином (VAMP). Синаптобревин переходит в активированное состояние и начинает выполнять роль своеобразного «крючка» или якоря. Этим якорем везикулы фиксируются к пресинаптической мембране в тех местах, где лежат особые белки – SNAP-25 и синтаксин-1. В последующем эти белки инициируют слияние мембраны везикул с мембраной аксона и выталкивают медиатор в синаптическую щель подобно поршню насоса. При прохождении потенциала действия через пресинаптическую мембрану одновременно опустошаются 2.000-3.000 везикул.
Схема 4. Передача сигнала в холинергическом синапсе. ХАТ – холинацетилтрансфераза, В1 – тиамин, Ach – ацетилхолин, М1-Хр – М1-холинорецепторы, АХЭ – ацетилхолинэстераза, ФлС – фосфолипаза С, PIP2 – фосфатидилинозитол бифосфат, IP3 – инозитол трифосфат, DAG – диацилглицерол, PkC – протеинкиназа С, Б – белок-фермент, Б-РО4 – фосфорилированная форма белка-фермента. Процесс выделения медиатора может быть нарушен под влиянием ботулотоксина (токсин бактерий Clostridium botulinum). Ботулотоксин вызывает протеолиз белков, участвующих в выделении медиатора (SNAP-25, синтаксин, синаптобревин). a-латротоксин – яд паука «черная вдова» связывается с белком SNAP-25 (нейрексином) и вызывает спонтанный массивный экзоцитоз ацетилхолина. 3. Развитие биологического ответа. В синаптической щели путем диффузии ацетилхолин поступает к постсинаптической мембране, где активирует холинорецепторы. При взаимодействии с Н-холинорецепторами происходит открытие натриевых каналов и на постсинаптической мембране генерируется потенциал действия. В том случае, если ацетилхолин активирует М-холинорецепторы, сигнал передается через систему G-белков на фосфолипазу С, ионные каналы К+ и Са2+ и все это приводит в конечном итоге к изменению поляризации мембраны, процессов фосфорилирования внутриклеточных белков. Помимо постсинаптической мембраны ацетилхолин может воздействовать на холинорецепторы пресинаптической мембраны (М1 и М2). При активации ацетилхолином М1-пресинаптического рецептора выделение медиатора усиливается (положительная обратная связь). Роль М2-холинорецепторов на пресинапетической мембране недостаточно ясна, полагают, что они могут тормозить секрецию медиатора. Развитие биологического ответа можно вызвать при введении лекарственных веществ, которые стимулируют холинорецепторы или предотвратить, если ввести средства, блокирующие эти рецептры. Повлиять на развитие эффекта можно и не затрагивая рецепторы, а воздействуя лишь на пострецепторные механизмы: · Токсин коклюшной палочки может активировать Gi-белок и снижать активность аденилатциклазы на затрагивая М-холинорецептор; · Токсин холерного вибриона может активировать Gs-белок и повышать активность аденилатциклазы; · Дитерпен форсколин из растения Coleus forskohlii способен непосредственно активировать аденилатциклазу в обход рецепторов и G-белков. 4. Окончание действия медиатора. Время существования ацетилхолина в синаптической щели составляет всего 1 мС, после чего он подвергается гидролизу до холина и остатка уксусной кислоты. Уксусная кислота быстро утилизируется в цикле Кребса. Холин в 1.000-10.000 раз менее активен, чем ацетилхолин, 50% его молекул подвергаются обратному захвату в аксон для ресинтеза ацетилхолина, остальная часть молекул включается в состав фосфолипидов. Гидролиз ацетилхолина осуществляет особый фермент – холинэстераза. В настоящее время известно 2 его изоформы: · Ацетилхолинэстераза (AChE) или истинная холинэстераза – осуществляет высокоспецифичный гидролиз ацетилхолина и локализуется на постсинаптической мембране холинергических синапсов. · Бутирилхолинэстераза (ButChE) или псевдохолинэстераза – осуществляет низкоспецифичный гидролиз эфиров. Локализуется в плазме крови и перисинаптическом пространстве. Сравнительная характеристика этих ферментов представлена в таблице 3. Таблица 3. Сравнительная характеристика холинэстераз. Дата добавления: 2014-11-24 | Просмотры: 1251 | Нарушение авторских прав |