|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Характеристика адренорецепторовАдренорецепторами называют молекулы клетки, которые реагируют на нейромедиатор норадреналин или гормон адреналин. Впервые мысль о возможном существовании нескольких типов адренорецепторов была выдвинута английским фармакологом Ahlquist в 1948 г. В настоящее время адренорецепторы подразделяют на 2 класса: · a-адренорецепторы – этот класс рецепторов опосредует сокращение гладких мышц под влиянием адреналина. Обнаружено 2 типа a-адренорецепторов (a1 и a2), каждый из которых имеет по крайней мере 3 подтипа. · b-адренорецепторы – этот класс рецепторов опосредует расслабление гладких мышц под влиянием адреналина. Обнаружено 3 типа b-адренорецепторов (b1, b2, b3). Все типы адренорецепторов являются семейством мембранных рецепторов, связанных с G-белками. Подробная их характеристика представлена в таблице 4. Таблица 4. Сравнительная харктеристика адренорецепторов.
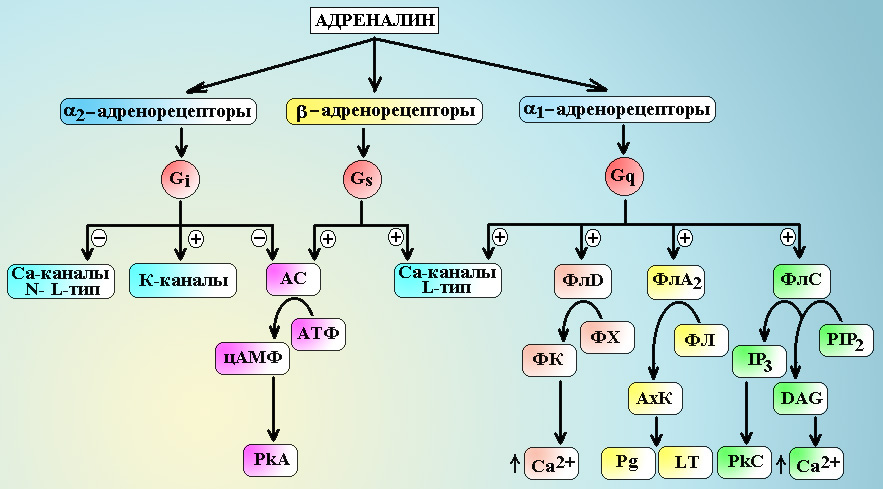
Примечание: A – адреналин, NA – норадреналин, Iso – изопреналин, МПС – мочеполовая система. a1-адренорецепторы посредством Gq-белка передают сигнал на несколько эффекторных систем: ] Фосфолипазу С, которая гидролизует фосфатидилинозитол бифосфат (PIP2) до инозитов трифосфата (IP3) и диацилглицерола (DAG). Молекулы IP3 вызывают выход ионов Са2+ из внутриклеточного депо и активируют зависимые от Са2+ ферменты (кальмодулин). DAG – обеспечивает активацию протеинкиназы С и фосфорилирование внутриклеточных белков, а также открывает Са2+-каналы мембраны. Под влиянием ионов Са2+ и активного кальмодулина происходит дефосфорилирование киназы легких цепей миозина и она переходи в активную нефосфорилированную форму, при этом начинают фосфорилироваться легкие цепи миозина и запускается процесс сокращения гладкомышечных клеток (см. схему 6.). ] Фосфолипазу А2, которая гидролизует фосфолипиды с выделение арахидоновой кислоты. В последующем арахидоновая кислота трансформируется в простагландины и лейкотриены. ] Фосфолипазу D, которая гидролизует фосфатидилхолин до фосфатидной кислоты. Молекулы фосфатидной кислоты вызывают выделение ионов Са2+ из депо, активируют АДФ-рибозилирующий фактор. ] Показана возможность активации G-белками Са2+-каналов клетки. a2-адренорецепторы посредством Gi-белка также передают сигнал на несколько эффекторных систем: ] Gi-белок снижает активность аденилатциклазы и уменьшает синтез цАМФ в клетке. В итоге, активность зависимых от цАМФ протеинкиназ падает. ] Через G0-белки тормозятся Са2+-каналы L- и N-типов. ] bg-субъединицы G-белка активируют К+-каналы мембраны. ] Относительно недавно было обнаружено, что bg-субъединицы Gi-белка могут стимулировать митоген-активирующие протеинкиназы (МАРК), которые обеспечивают процессы деления и размножения стволовых клеток.
Схема 5. Передача сигнала с адренорецепторов. АС – аденилатциклаза, PkA – протеинкиназа А, PkC – протеинкиназа С, ФлС – фосфолипаза С, ФлА2 – фосфолипаза А2, ФлD – фосфолипаза D, ФХ – фосфатидилхолин, ФЛ – фосфолипиды, ФК – фосфатидная кислота, АхК – арахидоновая кислота, PIP2 – фосфатидилинозитол бифосфат, IP3 – инозитол трифосфат, DAG – диацилглицерол, Pg – простагландины, LT – лейкотриены. b-адренорецепторы всех типов реализуют свое действие через Gs-белки. a-субъединицы этого белка активируют аденилатциклазу, которая обеспечивает синтез в клетке цАМФ из АТФ и активацию цАМФ зависимой протеинкиназы А. bg-субъединицы Gs-белка активируют Са2+-каналы L-типа и т.н. maxi-K+-каналы. Под влиянием цАМФ-зависимой протеинкиназы А происходит фосфорилирование киназы легких цепей миозина и она переходит в неактивную форму, не способную фосфорилировать легкие цепи миозина. Процесс фосфорилирования легких цепей прекращается и гладкомышечная клетка расслабляется. Дата добавления: 2014-11-24 | Просмотры: 1885 | Нарушение авторских прав |