|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
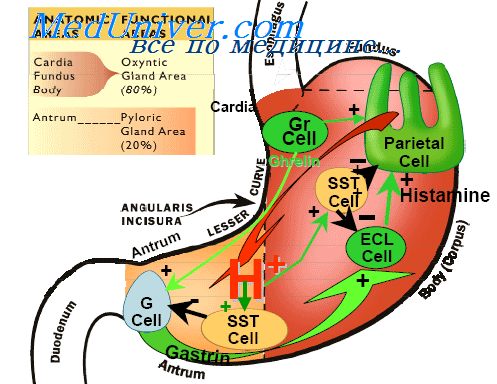
Нервная регуляция слюноотделенияНа рисунке показаны проводящие парасимпатические нервные пути, регулирующие слюноотделение. Видно, что слюнные железы контролируются в основном парасимпатическими нервными сигналами на всем протяжении от верхнего до нижнего ядер слюноотделения в стволе мозга. Ядра слюноотделения расположены приблизительно в месте соединения продолговатого мозга с мостом и возбуждаются вкусовыми и тактильными раздражителями от языка и других зон ротовой полости и глотки. Многие вкусовые стимулы, особенно кислый вкус (вызванный кислотой), приводят к обильной секреции слюны, часто в 8-20 раз превышающей базальный уровень секреции. Кроме того, обычные тактильные стимулы, такие как гладкие объекты во рту (например, галька), вызывают выраженную саливацию, в то же время шершавые объекты вызывают меньшую саливацию и в некоторых случаях даже блокируют слюноотделение. Слюноотделение также может быть вызвано или заторможено нервными сигналами, идущими в саливаторные ядра из более высоких отделов центральной нервной системы. Например, когда человек нюхает или ест любимую пищу, слюноотделение более выражено, чем при нелюбимой пище. Зона аппетита головного мозга, которая частично управляет этими эффектами, расположена ближе к парасимпатическим центрам переднего гипоталамуса. Ее функции в большей степени зависят от сигналов из вкусовой и обонятельной зон коры мозга или миндалевидного тела. Слюноотделение также возникает рефлекторно в ответ на раздражение желудка и верхнего отдела тонкой кишки, особенно когда проглочена раздражающая пища или когда человека тошнит от каких-либо желудочно-кишечных погрешностей. Проглатывание слюны помогает снизить раздражающий фактор в желудочно-кишечном тракте путем растворения или нейтрализации раздражающих веществ. Симпатическая стимуляция также может в небольшой степени усилить саливацию, но значительно меньше, чем парасимпатическая стимуляция. Симпатические нервы берут начало от верхнего шейного ганглия и идут вдоль стенок кровеносных сосудов в слюнные железы. Второстепенный фактор, который влияет на секрецию слюны, — это кровоснабжение желез, поскольку секреция всегда требует поступления достаточного количества питательных веществ из крови. Сигналы парасимпатических нервов, вызывающие обильную саливацию, также умеренно расширяют кровеносные сосуды. Помимо этого слюноотделение само непосредственно расширяет кровеносные сосуды, обеспечивая таким образом усиленное питание слюнной железы в соответствии с потребностями секретирующих клеток. Частично этот дополнительный сосудорасширяющий эффект обусловлен калликреином, который выделяют активированные клетки слюнных желез. Калликреин действует как фермент, расщепляющий один из белков крови (альфа2-глобулин) и образующий брадикинин, который является сильным вазодилататором. Секретом пищевода является только слизь, которая обеспечивает смазывание при глотании. Большая часть пищевода содержит множество простых слизистых желез. В гастральном и начальном (в меньшей степени) отделах пищевода также расположено большое количество сложных слизистых желез. Слизь, секретируемая этими железами в верхнем отделе пищевода, предотвращает повреждение слизистой вновь поступившей пищей, тогда как сложные слизистые железы, расположенные возле желудочно-пищеводного соединения, защищают стенку пищевода от раздражения кислыми желудочными соками, которые часто забрасываются из желудка обратно в нижний отдел пищевода. Несмотря на эту защиту, на гастральном конце пищевода нередко может образовываться пептическая язва. Помимо клеток, секретирующих слизь и покрывающих всю поверхность желудка, слизистая желудка имеет еще два важных типа тубулярных желез: фундалъные железы (также называемые собственными железами желудка) и пилорические железы. Фундальные железы (кислотопродуцирующие) выделяют соляную кислоту, пепсиноген, внутренний фактор и слизь. Пилорические железы выделяют главным образом слизь для защиты слизистой привратника от желудочной кислоты. Они также продуцируют гормон гастрин. Фундальные железы расположены на внутренней поверхности тела и дна желудка проксимального отдела, составляющего 80%. Пилорические железы находятся в антральном отделе дистальной части желудка, занимающего 20%. Типичная желудочная кислотопродуцирующая железа показана на рисунке. Она состоит из трех типов клеток:
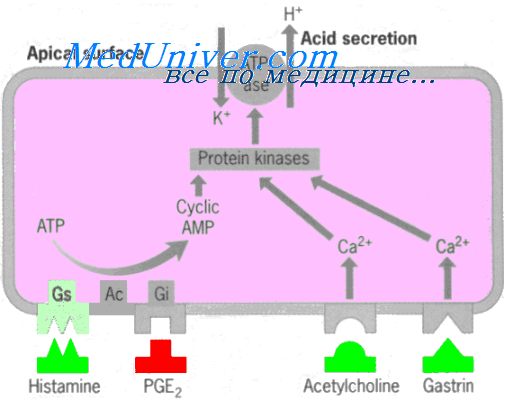
Основной механизм секреции соляной кислоты. Во время стимуляции париетальные клетки секретируют раствор кислоты, который содержит около 160 ммоль соляной кислоты на литр, что является практически изотоничным по отношению к жидкостям тела. рН этой кислоты составляет около 0,8, что свидетельствует о высокой кислотности. При таком рН концентрация ионов водорода приблизительно в 3 млн раза больше, чем в артериальной крови. Чтобы концентрировать ионы водорода в таком огромном количестве, требуется более 1500 калорий энергии на 1 л желудочного сока. На рисунке схематически показано функциональное строение париетальной клетки, которая содержит большие разветвляющиеся внутриклеточные канальцы. Соляная кислота образуется в ворсинкоподобных выступах внутри этих канальцев и затем проводится через канальцы в секреторный конец клетки. Были сделаны различные предположения относительно химических механизмов образования соляной кислоты. Один из них, включает следующие этапы. 1. Ион хлора активно транспортируется из цитоплазмы париетальной клетки в просвет канальца, а ионы натрия активно транспортируются из канальца в цитоплазму париетальной клетки. Эти два эффекта одновременно создают в канальце отрицательный потенциал от -40 до -70 мВ, что, в свою очередь, вызывает диффузию положительно заряженных ионов калия и небольшого количества ионов натрия из цитоплазмы клетки в каналец. Таким образом, КС1 и NaCl (в гораздо меньшей степени) входят в канальцы. 2. Вода цитоплазмы клеток диссоциирует на ионы водорода и гидроксилъные ионы. Ионы водорода затем активно секретируются в каналец в обмен на ионы калия; этот активный процесс обмена катализируется Н+/К+-АТФ-азой. К тому же ионы натрия активно реабсорбируются отдельным натриевым насосом. Таким образом, большая часть ионов калия и натрия, которые диффундируют внутрь канальца, реабсорбируются в цитоплазму клеток, а ионы водорода занимают их место в канальце, создавая в канальце концентрированный раствор соляной кислоты. Соляная кислота затем секретируется наружу через открытый конец канальца в просвет железы. 3. Вода поступает внутрь канальца благодаря осмосу из-за избытка ионов внутри канальца. Таким образом, конечный секрет канальца содержит воду, соляную кислоту в концентрации в пределах от 150 до 160 мэкв/л, КС1 — в концентрации 15 мэкв/л и малое количество. 4. В итоге диоксид углерода, образованный как в процессе обмена веществ в клетке, так и поступивший в клетку из крови, соединяется под действием карбоангидразы с гидроксильными ионами (пункт 2), образуя ионы бикарбоната. Затем эти ионы диффундируют из клеточной цитоплазмы во внеклеточную жидкость в обмен на ионы хлора, которые поступают в клетку из внеклеточной жидкости и позднее секретируются в каналец. Несколько малоразличаемых типов пепсиногена секретируются пептическими и слизистыми клетками желез желудка. Даже при таких условиях весь пепсиноген выполняет одинаковую функцию. Во время начальной стадии секреции пепсиноген не имеет переваривающей активности. Однако как только он вступает в контакт с соляной кислотой, он активируется, превращаясь в активный пепсин. Во время этого процесса молекула пепсиногена, имеющая молекулярную массу около 42500, расщепляется, образуя молекулу пепсина, имеющего молекулярную массу около 35000. Как активный протеолитический фермент пепсин работает только в сильно кислой среде (оптимальная рН от 1,8 до 3,5), а при рН около 5 он не имеет почти никакой протеолитической активности и за короткое время становится полностью неактивным. Соляная кислота, как и пепсин, также необходима для переваривания белка в желудке. Секреция внутреннего фактора. Внутренний фактору необходимый для всасывания витамина B12 в подвздошной кишке, секретируется париетальными клетками наряду с секрецией соляной кислоты. Когда париетальные клетки желудка погибают, что часто случается при хронических гастритах, у человека не только развивается ахлоргидрия (недостаток секреции соляной кислоты), но также часто возникает пернициозная анемия, обусловленная нарушением созревания красных клеток крови при отсутствии стимуляции костного мозга витамином B12.
Дата добавления: 2015-11-26 | Просмотры: 821 | Нарушение авторских прав |