|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
ПРОДОЛГОВАТЫЙ МОЗГ И ВАРОЛИЕВ МОСТПродолговатый мозг и варолиев мост вместе составляют задний мозг. Анатомически и функционально задний мозг связан со спинным мозгом, мозжечком и большими полушариями головного мозга. В нем замыкаются дуги многих сложнокоординирован-ных двигательных процессов.
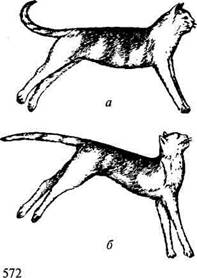
Сегментарное строение, четко выраженное в спинном мозге, стерто в продолговатом. Сюда вступают афферентные волокна от слуховых рецепторов, рецепторов полости рта, кожи лицевой части головы, сердца, крупных сосудов и легких, слизистых оболочек глаз, ряда рецепторов желудка, печени, поджелудочной железы, тонких кишок. Но в органах грудной и брюшной полости имеются также рецепторы, от которых афферентные волокна идут не в продолговатый, а в спинной мозг. Эфферентные волокна нейронов продолговатого мозга иннервируют все мышцы лица, сердце, бронхи, гортань, пищевод, желудок, поджелудочную железу, печень, кишечник. Иннервация внутренних органов осуществляется парасимпатическими волокнами совместно с симпатическими волокнами, лежащими в спинном мозге. Из области продолговатого мозга и варолиева моста выходят следующие черепно-мозговые нервы: тройничный, отводящий, лицевой, слуховой, языкоглоточный, блуждающий, добавочный, подъязычный. Продолговатый мозг играет большую роль в регуляции мышечного тонуса. Скелетные мышцы находятся в постоянном тонусе, за счет которого преодолевается действие силы тяжести и тело сохраняет равновесие. Тонус мышц имеет рефлекторную природу. Начало дуги этого рефлекса лежит в мышцах и сухожилиях. Если головной мозг вместе со средним мозгом и его красными ядрами удален, а продолговатый мозг сохранен и не отделен от спинного, то у млекопитающих развивается сильное, часами и сутками удерживаемое напряжение разгибательных мышц (рис. 13.13).
Конечности такого животного вытянуты, и согнуть их можно лишь с трудом; шейные мышцы, разгибающие голову, напряжены; позвоночник несколько выгнут дугой кверху. Таким образом, все мышцы, напряжение которых противостоит действию силы тяжести, оказываются после отделения продолговатого мозга от среднего в состоянии длительного тонического напряжения. Это Рис. 13.13. Децеребрационная ригидность: а — кошка с выключенными большими полушариями (путем перевязки сосудов); б— то же при одновременном выключении мозжечка резкое усиление тонуса разгибательных мышц носит название децеребрационной ригидности (децеребрация — удаление мозга выше продолговатого). Тонус разгибателей при децеребрационной ригидности превышает тонус сгибателей, но длительное напряжение сгибателей способствует тому, что конечности становятся как бы твердыми опорами и могут удерживать тяжесть тела. Децеребрационная ригидность не наступает, если средний мозг животного связан с продолговатым мозгом, а также если продолговатый отделен от спинного мозга. Отсюда следует, что в продолговатом мозге лежат нервные клетки, которые могут приводить в возбуждение спинальные нейроны, иннервирующие разгибатели. Если продолговатый мозг связан со средним мозгом, ригидность также не наступает, т. е. от среднего мозга исходят влияния, угнетающие стимуляцию разгибательных рефлексов со стороны продолговатого мозга. Заключительный этап в возникновении непомерного тонуса связан с рефлекторной деятельностью ядра вестибулярного нерва. Волокна от рецепторов вестибулярного аппарата в продолговатом мозгу контактируют с массой клеток, образующих так называемое вестибулярное ядро. Оперативное удаление вестибулярного ядра и особенно его части, известной как ядро Дейтерса, резко ослабляет децеребрационную ригидность. Разрушение вместе с ядром Дейтерса ретикулярной формации продолговатого мозга совершенно ее прекращает. Для развития децеребрационной ригидности необходимо поступление в ЦНС импульсов от рецепторов разгибательных мышц. Инъекция в эти мышцы раствора новокаина или перерезка соответствующих задних корешков снимает тонус мышц-разгибателей. Следовательно, децеребрационную ригидность надо рассматривать как результат усиления рефлексов, осуществляемых совместно спинным и продолговатым мозгом в ответ на импульсы, поступающие как от рецепторов мышц-разгибателей, растягиваемых под действием силы тяжести, так и от рецепторов лабиринта. Эти рефлексы в норме всегда уравновешиваются воздействиями, рефлекторно возникающими в среднем мозге, мозжечке, а у высших млекопитающих главным образом в коре больших полушарий. Роль продолговатого мозга в осуществлении децеребрационной ригидности обусловлена постоянным поступлением импульсов от важнейших рецепторов дыхательной и кровеносной систем, а также с лабиринтов, вследствие этого он постоянно воздействует на рефлекторные центры спинного мозга, повышая их возбудимость. При отделении спинного мозга от продолговатого возбудимость нервного центра никогда не достигает той степени, при которой тонус мышц-разгибателей в ответ на импульсы от их рецепторов рефлекторно усиливался бы до степени, характеризующей децеребрационную ригидность. Децеребрационная ригидность — это состояние, конечно, совершенно нефизиологичное, но по ней можно судить о взаимоотношениях между разными образованиями ЦНС. Надо заметить, что механизм этого процесса еще не совсем ясен. Это состояние не всегда наступает после децеребрации, иногда оно выражено очень резко, а иногда почти не проявляется. Продолговатый мозг участвует в регуляции проводниковых рефлексов и в осуществлении таких актов, как ходьба и стояние, для которых необходима интегративная деятельность ЦНС. Буль-барное животное, у которого сохранены лишь спинной и продолговатый мозг, способно к более сложным рефлекторным процессам, чем спинальное, так как все основные функции в более полной мере объединены и скоординированы. Варолиев мост является центральной частью заднего мозга. Он имеет вид толстого белого вала. Серое вещество образуют ядра черепно-мозговых нервов (V—VIII пары), ядра ретикулярной формации и собственные ядра моста. Белое вещество состоит из продольных и поперечных пучков нервных волокон. Продольные волокна идут по нижней поверхности моста в продолговатый мозг, образуя в нем пирамиды. Сверху они составляют восходящие и нисходящие пути двусторонних связей моста. Поперечные волокна направляются в ножки мозжечка. Чувствительные волокна черепно-мозговых нервов проводят сигналы от вкусовых рецепторов языка, рецепторов кожи и мышц головы, глаз и зубов. Эфферентные волокна обеспечивают мимику. Таким образом, рефлекторная деятельность варолиева моста дополняет защитные и пищевые рефлексы, осуществляемые продолговатым мозгом. СРЕДНИЙ МОЗГ Средний мозг, так же как продолговатый мозг и варолиев мост, относится к стволовым структурам. Он представляет собой относительно небольшую часть мозгового ствола. Через средний мозг проходят все восходящие пути, передающие импульсы к таламусу, большим полушариям и мозжечку, а также нисходящие пути, проводящие импульсы к продолговатому и спинному мозгу. В среднем мозге, так же как и в продолговатом, расположены нейроны ретикулярной формации. Анатомически средний мозг состоит из двух основных частей: дорсального отдела, который представляет собой крышку мозга, или четверохолмие, и базального, который представлен его ножками. Каждая из ножек, в свою очередь, состоит из покрышки, черной субстанции и основания. Покрышка — это продолжение дорсальной части моста, в которой находятся красное ядро и ядро блокового и глазодвигательного нервов (рис. 13.14).
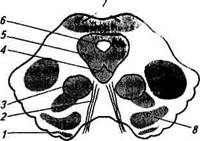
Рис. 13.14. Разрез через средний мозг на уровне ядер глазодвигательного нерва: / — ножки мозга; 2— глазодвигательный нерв; 3— красное ядро; 4— ядро глазодвигательного нерва; 5— центральное серое вещество; б—водопровод мозга (сильвиев водопровод); 7— переднее двухолмие; 8— черная субстанция Рефлекторные функции ядер среднего моз-г а. Передние бугры четверохолмия являются первичными зрительными центрами, при участии которых осуществляются некоторые рефлексы в ответ на световые раздражения. К таким рефлексам относятся зрительные ориентировочные рефлексы, проявляющиеся в том, что животное, лишенное больших полушарий, но с сохраненным средним мозгом, реагирует на световое раздражение движением глаз и поворотом головы. Рефлекторные движения глаз происходят благодаря поступлению к глазным мышцам импульсов от ядер глазодвигательного и блокового нервов. Передние бугры четверохолмия принимают участие в осуществлении зрачкового рефлекса, аккомодации глаза, а также в конвергенции — сведении зрительных осей. Задние бугры четверохолмия представляют собой первичные слуховые центры. При их участии осуществляются ориентировочные слуховые рефлексы. При ориентировочных рефлексах у животных с целым средним мозгом наблюдаются и вегетативные рефлексы: изменение ритма сердечной деятельности, артериального давления. Ядра четверохолмия обеспечивают сторожевой рефлекс, значение которого состоит в подготовке реакции организма на внезапное раздражение. При этом происходит перераспределение мышечного тонуса: усиление тонуса сгибателей, что способствует быстрой ответной реакции животного. В ножках мозга идут проводящие пути как от нижележащих центров в высшие центры головного мозга, так и от высших центров в спинной и продолговатый мозг. Эти же проводящие пути образуют ножки мозжечка и пучки волокон между скоплениями нервных клеток. Черная субстанция отвечает за координацию актов глотания и жевания, а также участвует в регуляции пластического тонуса; имеет значение при выполнении пальцами мелких движений, требующих большой точности. Поэтому черная субстанция лучше развита у приматов. Красное ядро — крупное скопление серого вещества среднего мозга, состоит из крупных и мелких клеток. Оно связано проводя-
Тонические рефлексы ствола мозга. Важнейшая функция ствола мозга, в частности продолговатого и среднего мозга, состоит в перераспределении мышечного тонуса в зависимости от положения тела в пространстве. Перераспределение тонуса осуществляется рефлекторным путем и обеспечивает сохранение равновесия тела. Всю совокупность тонических рефлексов голландский ученый Р. Магнус разделил на две большие группы: статические рефлексы; статокинетические рефлексы. Статические рефлексы. Выражены при спокойном стоянии, лежании и сидении в различных позах, они обусловливают определенное положение тела в пространстве. Такие рефлексы делятся на две большие группы. Первая группа обеспечивает определенное положение или позу тела — это рефлексы положения, или по-зотонические рефлексы. Вторая же способствует возвращению тела из неестественного положения в нормальное — это установочные, или выпрямительные, рефлексы. Позотонические рефлексы включают в себя лабиринтные и шейные. Лабиринтные рефлексы, для осуществления которых достаточно наличия продолговатого и спинного мозга, обеспечивают фиксирование конечностей в определенном положении. В эксперименте на шею децеребрированного животного надевали гипсовую повязку, чтобы голова сохраняла постоянное положение по отношению к туловищу. При этом не происходит раздражение проприорецепторов шеи. Затем животное вращали вокруг оси, условно проведенной через оба ^иска. В положении, когда животное было спиной вниз, тонус разгибателей становился максимальным и конечности вытягивались, а если же оно оказывалось спиной вверх, то тонус этих мышц снижался и конечности сгибались. На тонус всех мышц помимо лабиринтных влияют также шейные рефлексы. Влияние шейных рефлексов сказывается в том, что при поворотах головы по отношению к туловищу справа налево или слева направо тонус разгибателей максимален на той стороне тела, к которой обращено темя. Если же поворачивать голову вокруг оси, условно проведенной через оба слуховых прохода, то при наклоне головы вниз тонус разгибателей увеличивается в зад- них и снижается в передних конечностях. При запрокидывании головы назад, наоборот, тонус разгибателей передних конечностей увеличивается, а задние конечности сгибаются. Рефлексы с лабиринтов на шейные мышцы. Шейные и лабиринтные тонические рефлексы осуществляются в связи друг с другом, так как импульсы с лабиринтов влияют на тонус шейных мышц. Максимум тонуса шейных мышц наблюдается при тех положениях головы, когда импульсы с лабиринтов ведут к рефлекторному усилению тонуса разгибателей конечностей. Импульсы с каждого лабиринта влияют преимущественно на шейные мышцы противоположной стороны (поэтому при разрушении лабиринта голова животного повернута в сторону повреждения). Выпрямительные рефлексы осуществляются средним мозгом и, следовательно, отсутствуют у бульбарных животных. В осуществлении этих рефлексов участвуют рецепторы лабиринтов, шейных мышц и кожи. Если мезенцефальное животное положить на бок, оно поднимает голову и устанавливает ее в естественное положение теменем кверху. Это происходит в результате возбуждения рецепторов вестибулярного аппарата. Но даже при разрушении вестибулярного аппарата голова выпрямляется в том случае, если тело лежит на боку на какой-нибудь твердой поверхности. В этом случае в ответ на одностороннее раздражение кожи происходит рефлекторное выпрямление головы. Если на это же животное положить сверху доску с небольшим грузом, то происходит симметричное двустороннее раздражение кожных рецепторов и голова снова падает вниз. Если доску убрать, то голова рефлекторно снова поднимается. Подъем головы —это только первая фаза выпрямительных рефлексов. Вторая фаза заключается в рефлекторном выпрямлении туловища, которое следует за подъемом головы. Этот рефлекс возникает от раздражения проприорецепторов мышц шеи и рецепторов кожи туловища. Статокинетические рефлексы. Начинаются с одного рецептивного поля — с рецепторов полукружных каналов внутреннего уха. Возникают при угловом или линейном ускорении. При вращении животного наблюдается нистагм головы, т. е. медленный поворот в сторону, противоположную вращению, и быстрый возврат в исходное положение. Глаза реагируют на вращение аналогичным образом — глазной нистагм. Осуществление статокинетичес-ких рефлексов возможно только при участии ядер среднего мозга. Дата добавления: 2015-12-15 | Просмотры: 1158 | Нарушение авторских прав |
 щими путями с мозжечком и ядрами полосатого тела, а также со §пинным мозгом (руброспинальный путь). Красные ядра с окружающей их ретикулярной формацией, несомненно, участвуют в регуляции мышечного тонуса. Можно сравнить поведение деце-|ебрированного животного, у которого со спинным и продолговатым мозгом сохранилась связь лишь посредством варолиева моста, мозжечка и нижнего участка среднего мозга без области красных ядер, с поведением так называемого таламического животного, у которого удалены кора мозга и подкорковые узлы, но средний мозг с его красными ядрами и зрительные бугры сохранены и остаются в связи с продолговатым мозгом.
щими путями с мозжечком и ядрами полосатого тела, а также со §пинным мозгом (руброспинальный путь). Красные ядра с окружающей их ретикулярной формацией, несомненно, участвуют в регуляции мышечного тонуса. Можно сравнить поведение деце-|ебрированного животного, у которого со спинным и продолговатым мозгом сохранилась связь лишь посредством варолиева моста, мозжечка и нижнего участка среднего мозга без области красных ядер, с поведением так называемого таламического животного, у которого удалены кора мозга и подкорковые узлы, но средний мозг с его красными ядрами и зрительные бугры сохранены и остаются в связи с продолговатым мозгом.