|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
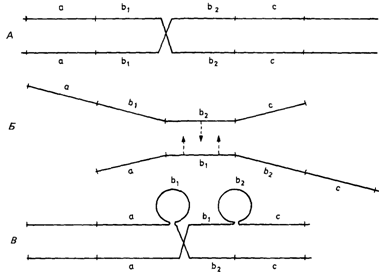
Неравный кроссинговерОткрытие неравного кроссинговера. В первые годы работы с дрозофилой некоторые авторы обратили внимание на то, что мутация Bar (Х-сцепленный доминантный признак) иногда ревертировала к нормальному фенотипу. Гомозиготы по этому аллелю давали потомство, несущее новый аллель, позже названный «двойной Bar», с еще более выраженным эффектом. Стертевант (1925) [904] показал, что такое необычное поведение было следствием не точковых мутаций, а неравного кроссинговера, приводящего к появлению хромосомы с двумя локусами Bar (двойной Bar) и одновременно хромосомы вовсе без этого локуса. Когда методика работы с гигантскими хромосомами слюнных желез дрозофилы позволила визуально проверять генетические гипотезы, Бриджесу (1936) [588] удалось показать, что простая доминантная мутация Bar была вызвана дупликацией хромосомного диска. Реверсия соответствует недушшцированному состоянию, тогда как двойной Bar вызывается трипликацией этого диска. Как реверсия, так и трипликация порождаются одним-единственным событием неравного кроссинговера. Тем не менее Бриджес не сформулировал четко причину этого события, по его мнению, она заключается в ошибочном спаривании «структурно-гомологичных», а не «позиционно-гомологичных» хромосомных сайтов (рис. 3.43, 2.97). Неравный кроссинговер в генетике человека. Гаптоглобин- транспортный белок для гемоглобина, содержащийся в сыворотке крови [584а]. Наиболее распространенные в популяции аллели обозначаются HP1F, HP1S и HP2. В 1962 г. было обнаружено [884], что аллель HP2, судя по первичной структуре соответствующей полипептидной цепи, почти вдвое длиннее каждого из двух аллелей HP1F и HP1S. В НР2-цепи аминокислотная последовательность НР1цепи повторяется почти полностью. Авторы сделали вывод о том, что аллель HP2 возник в результате генной дупликации. Кроме того, они предсказали, что существует относительно высокая вероятность
228 3. Формальная генетика человека
повторного неравного кроссинговера между НР2-аллелями, что должно привести к появлению, с одной стороны, аллеля, подобного HP1, а с другой – аллеля, содержащего почти утроенную генетическую информацию. Последующие кроссинговеры между такими аллелями могут привести к образованию еще более протяженных генов и, следовательно, к полиморфизму в популяции по длине аллелей. Имеется существенное различие между первым уникальным событием, в результате которого образуется ген почти вдвое длиннее обычного (например, HP2) из единичного гена HP1, и неравным, но гомологичным кроссинговером, который становится возможным, как только в популяции появится дуплицированный аллель [748]. Первое событие. Пусть имеется пара гомологичных хромосом, причем оба партнера содержат протяженные идентичные нуклеотидные последовательности. В норме эти партнеры спариваются в мейозе, и неравный кроссинговер не может произойти. Чтобы вызвать ошибочное спаривание и тем самым неравный кроссинговер, необходима исходная дупликация по крайней мере одного гена. В цитогенетике известны механизмы такой дупликации, простейший из которых - два разрыва в немного смещенных (один относительно другого) сайтах в конъюгирующих гомологичных хроматидах во время мейоза и последующее перекрестное воссоединение. Другой механизм – это ошибочное спаривание вследствие гомологии коротких нуклеотидных последовательностей в негомологичных положениях. По современным представлениям структура последовательностей ДНК между транскрибируемыми участками генов (разд. 2.3) допускает много разных возмож- 3. Формальная генетика человека 229
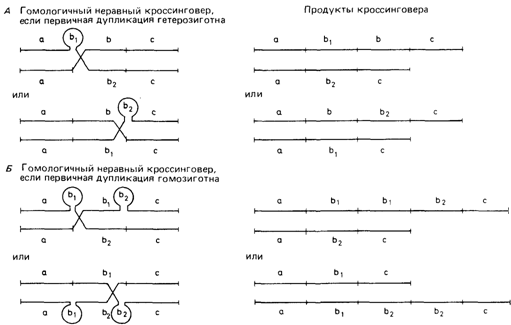
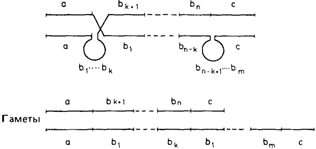
ностей для такого ошибочного спаривания. Если сайты разрыва отстоят только на длину одного структурного гена, то результатом этого события будут четыре гаметы: две, вовсе не содержащие данный ген, и две другие, содержащие его в дуплицированном виде (рис. 3.44). Гаметы, содержащие делецию, с высокой вероятностью могут утратиться вследствие гибели эмбриона. С другой стороны, гамета с дупликацией приведет к появлению диплоидного индивида, у которого в мейозе возможно ошибочное спаривание гомологичных цистронов и, следовательно, неравный кроссинговер. Последствия неравного кроссинговера. Последствия показаны на рис. 3.44. Пока дупликация остается гетерозиготной, все гаметы будут содержать либо одну, либо две копии дуплицированного гена. Однако, когда дупликация становится гомозиготной, возникают другие типы гамет. В результате неравного кроссинговера могут образоваться, с одной стороны, гаметы с единственной копией, а с другой - гаметы, содержащие три, а в последующих поколениях и большее число копий данного гена (рис. 3.44, 3.45). Если вероятность неравного кроссинговера не слишком мала, то довольно быстро создается высокая изменчивость по числу гомологичных хромосомных сегментов, которые, оставаясь сходными по первичной структуре, различаются по положению. Если отбор благоприятствует определенному числу таких хромосомных сегментов, то вскоре это число станет наиболее распространенным. Затухание отбора приведет к увеличению изменчивости в обоих направлениях. Постепенно увеличится доля идивидов как с очень высоким, так и с небольшим числом таких генов [748]. Другой генетический механизм, сходный в некоторых аспектах с неравным кроссинговером, - это генная конверсия, в результате которой образуются нереципрокные продукты (разд. 2.3, рис. 2.97). Возможное значение в генетике человека. Как уже упоминалось, гаптоглобиновый аллель HP2 представляет собой почти полностью дуплицированный HP1-аллель. В этом случае неравный кроссинговер, как ожидается, приводит иногда к появлению аллелей, содержащих утроенную информацию. Такие аллели на самом деле иногда наблюдались. Они известны как аллели типа Джонсона [883]. Другим примером могут служить тесно сцепленные глобиновые β- и δ-цистроны (разд. 4.3). В этом случае неравный кроссинговер может породить мутанты типа Лепоре (рис. 4.51), а также Х-сцешгенные гены цветоощущения [825а]. Кроме того, имеется много примеров умеренно и высокоповторяющихся последовательностей ДНК, внутри которых возможен неравный кроссинговер. Гены рибосомных РНК, локализованные внутри районов ядрышкового организатора, имеют около 300-400 идентичных копий с заметной вариацией. На первый взгляд такая ситуация обеспечивает наилучшие условия для неравного кроссинговера. Однако эти гены локализованы близко к центромере акроцентрических хромосом, где кроссинговер вряд ли происходит.
230 3. Формальная генетика человека
Остаются нерешенными вопросы, может ли вообще происходить кроссинговер в дистальных районах акроцентрических хромосом и не является ли акроцентрическая локализация защитным механизмом против отклонения от оптимального количества рРНК-генов за счет неравного кроссинговера? Гены, кодирующие иммуноглобулины (разд. 4.4), также являются повторяющимися последовательностями ДНК. Чем больше мы узнаем о функциональном значении повторяющихся последовательностей ДНК, тем лучше будем понимать роль неравного кроссинговера. Внутрихромосомный неравный кроссинговер. У структурно-гомологичных (но не позиционно-гомологичных) генов, таких, как найденные в мультигенных семействах (разд. 2.3.3.8), неравный кроссинговер происходит не только между гомологичными хромосомами, но также между сестринскими хроматидами (внутрихромосомный неравный кроссинговер). Теоретические рассуждения показывают, что этот процесс мог сыграть определенную роль в молекулярной эволюции [1941]. Дата добавления: 2015-12-16 | Просмотры: 1815 | Нарушение авторских прав |