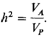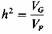|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Концепция наследуемостиКонцепция наследуемости широко применяется в количественной генетике. Градации изучаемого признака, выраженные в метрических единицах, могут быть названы «значениями». Значение, измеренное у данного индивида, является его фенотипическим значением. Для большинства биологических признаков это фенотипическое значение определяется как генетическими, так и средовыми факторами. Среда рассматривается в широком смысле, т. е. охватывает все негенетические факторы, оказывающие влияние на фенотип (Falconer [63] использует термин средовое отклонение): Р = G + Е, где Р — фенотипическое значение, G-генотипическое значение и Е - средовое значение. Фенотипические значения всех индивидов в популяции имеют среднее значение и дисперсию, которая измеряет вариацию вокруг среднего. Дисперсия отличается от других мер изменчивости одним математическим свойством: различные дисперсии можно складывать, получая величину общей дисперсии, и наоборот, общую фенотипическую дисперсию VP можно подразделить на компоненты, такие, как генотипическая дисперсия VG и средовая VE: VP = VG + VE. 3. Формальная генетика человека 243
Однако это правило для дисперсии применимо, только если генотипические и средовые значения независимы друг от друга, т. е. когда они некоррелированы. Если между ними имеется корреляция, то следует добавить ковариацию G и Е: VP=VG+VE + 2CovGE Давайте возьмем один пример из той области генетики, в которой впервые была введена эта концепция - сельскохозяйственные исследования (Falconer [63]). В молочном животноводстве практика кормления коров соответствует их молочной продуктивности: коровам, от которых получают больше молока, дают больше корма. Человеческое общество часто поступает точно так же в отношении своих членов. Этой темы мы коснемся в разделе, посвященном генетике поведения. Другое предположение состоит в том, что определенные средовые различия имеют одинаковое влияние на разные генотипы. Когда это не так, то существует взаимодействие между генотипом и средой, которое дает дополнительную компоненту V 1в общей дисперсии. Даже для экспериментальных животных эта компонента может быть измерена только при особых условиях. Генотипическая дисперсия V G может быть подразделена на несколько компонент: аддитивную компоненту (VA) и компоненту (VD), измеряющую отклонение от ожидаемого значения в аддитивной модели, которое возникает вследствие доминирования и эпистаза. В доминантную дисперсию вносят вклад гетерозиготы (Аа), которые не занимают строго промежуточного положения между соответствующими гомозиготами (аа и АА). Вклад в дисперсию, осуществляемый эпистазом, относится к действию тех генов, которые влияют на экспрессию других генов. Следовательно, концепция аддитивной дисперсии не подразумевает допущения чисто аддитивного действия генов. Даже действие генов, обнаруживающих доминирование и эпистаз, проявляет тенденцию иметь аддитивную компоненту. Таким образом, в общей фенотипической дисперсии можно выделить следующие компоненты:
Здесь введена новая компонента (VM), которая относится к ваьированию измерений одного и того же признака в разное время. Это могут быть результаты разных опытов в разные дни, или ошибки измерений при тестироваии одного и того же образца крови, или различия в повторных обследованиях одного и того же индивида. Если все эти переменные известны, то их тоже можно включить в вычисления. Далее, можно предложить, что ковариация между наследственностью и средой (CovGE) и дисперсия взаимодействия (V 1 ) равны нулю и их можно опустить. Для простоты мы будем полагать также, что компонентой воспроизводимости (VM) можно пренебречь (мы рассмотрим ее позже в разделе, посвященном близнецовому методу, см. прил. 6). Для удобства полезно ввести новое понятие-понятие наследуемости, которое определяется так:
Коэффициент наследуемости принимает значения от 0 до 1 (или от 0 до 100%) и выражает вклад аддитивных генетических факторов в изучаемый фенотип. Другими словами, можно сказать, что наследуемость-это популяционный статистический параметр, который выражает (аддитивный) генетический вклад в изучаемый признак в процентах. Низкое значение наследуемости подразумевает малый вклад аддитивно действующих генов в признак, тогда как высокое значение - большой вклад. Эта концепция была развита в связи с селекцией растений и животных, в частности, таких экономически полезных признаков, как молочная продуктивность коров и яйцено- 244 3. Формальная генетика человека скость кур. Для этих целей наиболее важной была аддитивная чась генетической изменчивости. Любая другая генетическая компонента, такая, например, как доминирование, снижает точность предсказания значений признака у потомства по значениям признака у родителей. Что касается человека, то мы больше интересуемся общей генетической изменчивостью: является ли она вся аддитивной или нет. В генетике человека наследуемость, как она была определена выше, часто называют «наследуемостью в узком смысле» и дополняют другим понятием:
где VG и VP относятся к общей генотипической и фенотипической дисперсии соответственно. Это определение известно как «наследуемость в широком смысле» или степень генетической детерминации. Существует соотношение между наследуемостью в узком смысле (hN2) и теоретическим коэффициентом корреляции между родственниками, как он определялся и вычислялся выше (3.5). Для наиболее важных степеней родства принимаются следующие формулы:
Свойства h 2. Для интерпретации биологического смысла оценки наследуемости необходимо тщательно рассмотреть ее свойства. 1. Наследуемость - это отношение. Оно изменяется, если изменяется числитель или знаменатель, h2 увеличивается, когда увеличивается числитель (генотипическая VG или аддитивная VA дисперсии) или уменьшается средовая дисперсия VE. 2. Оценка дисперсии основывается на теоретических корреляциях между родственниками. Эти корреляции справедливы только в предположении случайного скрещивания. Ассортативное скрещивание приводит к другим корреляциям, и если оно не учитывается, то это порождает систематические ошибки в оценке h 2. Корреляции при ассортативном скрещивании впервые были вычислены Фишером [664], а также Кавалли-Сфорца и Бодмером [36]: более полная модель описана Вильсон [955; 956]. Эти корреляции можно использовать для коррекции оценок h 2 3. Оценка h 2 возможна только в том случае, когда делается предположение, что ковариация и взаимодействие между генетическим значением и средовым отклонением равны нулю. Фалконер попытался избежать этой дилеммы для ковариации, сделав следующее допущение: если проявление генетической конституции индивида модифицируется средовыми условиями, «улучшая» или «ухудшая» его фенотип, то этот вклад можно включить в генотипическое значение как его часть. Формально это правильно, даже если такое допущение усложняет проблему взаимоотношений генотипа и фенотипа. В селекции животных это допущение может быть полезным, но в генетике человека его применение недопустимо. Еще большие трудности возникают, когда эти концепции применяются для интерпретации значений наследуемости, получаемых на основе близнецовых данных (приложение 6). Компонента в разложении дисперсии, связанная с взаимодействием, приводит к другой трудности в интерпретации, которая до сих пор еще не преодолена. Дата добавления: 2015-12-16 | Просмотры: 716 | Нарушение авторских прав |