|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Концепция генетического грузаТеория Определение общего числа рецессивных генов в популяциях человека [1856; 1871]. Гомозиготы, особенно по редким заболеваниям, среди детей от кровнородственных браков встречаются чаще, чем в популяции в целом. Исходя из этого, можно определить число таких рецессивных генов у каждого индивида в популяции. Например, пусть вероятность того, что ген, случайно выбранный у индивида, идентичен по происхождению одному из двух аллелей того же локуса его брата (сестры), равна 1/2. Если один из сибсов в браке брат—сестра несет ген, который в гомозиготном состоянии вызывает рецессивное заболевание, то другой сибс имеет этот ген с вероятностью 1/2, а риск заболевания для каждого ребенка этой пары равен 1/4. Следовательно, вероятность того, что хотя бы один ребенок от этого брака окажется больным, равна l–(3/4) s, где s- число детей в браке. Таким образом, путем анализа частоты рецессивных болезней в потомстве некоторого числа браков между братьями и сестрами (при условии, что отбор по каким-либо другим признакам отсутствует) можно определить среднее число особей в популяции, имеющих этот рецессивный ген. Те же самые рассуждения справедливы для браков отец—дочь. Однако браки между столь близкими родственниками встречаются крайне редко и запрещены законом. Кроме того, индивидов, вступающих в по- 350 6. Популяционная генетика добные браки, никак нельзя считать репрезентативной выборкой из популяции (см. раздел 6.3.2.4). Интуитивные предпосылки: наш груз мутаций. Известного генетика Г. Мёллера с юных лет занимала мысль о том, что для человека как вида существует опасность биологического вырождения. Он считал, что рано или поздно человечество погрузится в пучину страданий от болезней и умственных расстройств. В начале века эти опасения разделяли многие ученые; именно они явились причиной появления работ Ф. Гальтона и евгенического движения. Аргументы Мёллера были подробно изложены в его статье «Наш груз мутаций» (1950) [1835]. Наиболее важные положения этой статьи можно сформулировать следующим образом: а) большинство зигот человека в результате мутаций погибает или утрачивает способность к делению; б) общая скорость мутирования на индивид, т.е. общее число новых мутаций, содержащихся в тех двух зародышевых клетках, от которых он происходит, составляет одну мутацию на 2—10 зародышевых клеток; в) каждый индивид гетерозиготен по нескольким генам, летальным в гомозиготном состоянии; эти гены обычно даже в гетерозиготном состоянии оказывают вредное действие; г) из-за ослабления интенсивности естественного отбора число вредных генов в популяциях человека угрожающе растет; их частота может превысить критический уровень, после чего генетическая система человека разрушится и человек как вид исчезнет; д) эта опасность обостряется в связи с увеличением воздействия ионизирующей радиации; е) необходимо противостоять опасным тенденциям путем искусственной регуляции размножения человека. С тех пор как Мёллер сформулировал эти положения, наши знания о генетике человека пополнились и на некоторые из поставленных им вопросов мы получили довольно точный ответ [1886]. Выделим один из них, а именно утверждение, что каждый человек гетерозиготен по нескольким генам, летальным в гомозиготном и неблагоприятным даже в гетерозиготном состоянии. Влияние изменчивости на приспособленность. Более формализованная и рациональная концепция была разработана Холдейном в нескольких статьях, особенно в одной, названной «Влияние изменчивости на приспособленность» [1775; 1780]. Холдейн определил приспособленность генотипа как среднее число потомков, оставляемое особью этого генотипа, и отметил, что средняя приспособленность вида почти всегда близка к единице, так как в противном случае размер популяции будет очень быстро увеличиваться. Однако у любого вида приспособленность некоторых генотипов ниже 1 и падает до нуля в случае летальных генов и генов, вызывающих полную стерильность. Следовательно, приспособленность «стандартного» генотипа, не содержащего неблагоприятных генов, должна превышать единицу. Совершенно ясно, что в состоянии равновесия гены с вредным эффектом отметаются естественным отбором точно с такой же скоростью, с какой они возникают в результате мутирования. Летален ли ген или почти безвреден, значения не имеет. В первом случае отбором отметается каждая особь, имеющая такой ген, или, если этот ген рецессивный, каждая особь, гомозиготная по этому гену. Во втором случае жизнеспособность или плодовитость несущих данный ген особей может уменьшиться только на одну тысячную. Однако в обоих случаях падение приспособленности вида зависит только от скорости мутирования, а не от влияния гена на приспособленность несущей его особи. Внимательный читатель, возможно, уже понял, что это утверждение является обобщением аргументов Холдейна, обосновывающих косвенный метод оценки частоты мутирования у человека (разд. 5.1.3.1). Проведя предварительный анализ общей скорости мутирования у дрозофилы, Холдейн продолжает: «Это та цена, которую вид платит за изменчивость, являющуюся, вероятно, необходимым условием эволюции». 6. Популяционная генетика 351 В более поздней работе [1780] Холдейн оценил, насколько должна уменьшиться приспособленность, чтобы в ходе эволюции в результате действия естественного отбора произошло замещение адаптивного гена. Концепция генетического груза была использована для оценки мутирования в популяциях человека Мортоном в его работе: «Определение мутационного груза у человека на основе данных по кровнородственным бракам» [1827]. Определение генетического груза [1827]. Мортон, Кроу и Мёллер [1827] различают общий генетический груз, обусловленный вредными мутациями, присутствующими в геноме человека, и выявляемый (expressed) генетический груз; и тот и другой выражаются в летальных эквивалентах. Летальный эквивалент - это такое число мутаций, которое, будучи распределено среди нескольких особей, в среднем приводит к одному летальному исходу по генетическим причинам. Например, летальному эквиваленту соответствует одна летальная мутация, которая обусловливает гибель особи во всех случаях, или две мутации, каждая из которых приводит к гибели в 50% случаев. Общий груз на гамету определяется как среднее число летальных эквивалентов на такую зиготу, которая образуется путем удвоения всех хромосом гаметы. Выявляемый груз на гамету - это среднее число летальных эквивалентов, которое проявилось бы в том случае, если бы эта гамета образовала зиготу при соединении с другой гаметой в соответствии с системой скрещивания, преобладающей в данной популяции. Общий генетический груз можно оценить следующим образом. Рассмотрим один генный локус. Вероятность выживания данной зиготы, несмотря на вредный эффект мутаций по этому локусу, равна
Здесь s = вероятность гибели зиготы, гомозиготной по данной мутации; й-степень доминантности этой мутации (h = 0, если ген полностью рецессивен, h = 1, если ген приводит к гибели в гомозиготном и гетерозиготном состоянии с одинаковой частотой); F-коэффициент инбридинга. Второе допущение предполагает независимое действие генетических и средовых причин, приводящих к гибели. При этом условии доля выживших зигот оценивается следующим образом:
Здесь Xj - вероятность гибели в результате влияния какого-либо фактора среды. Произведение включает все х, и qs (частоты вредных мутаций). Предполагается, что число этих мутаций и число факторов среды xt велико, а отдельные значения вероятностей малы. Поэтому это выражение можно аппроксимировать следующим образом:
Это в свою очередь аппроксимируется как S = e-('4+BF) или - In S = А + BF, (6.16) где
Суммирование производится по всем факторам среды и соответственно по всем локусам с мутантными аллелями. В случайно скрещивающейся популяции (F = 0) выявляемый генетический груз вместе со средовым грузом равен А. Величина В, с другой стороны, измеряет скрытый генетический груз, который проявляется только в случае полной гомозиготности (F = 1). Общий генетический груз равен В и А можно оценить, используя взвешенные коэффициенты регрессии lns (s – доля выживающих особей) на F. Учитывая низкую степень инбридинга, наблюдающуюся обычно в популяциях человека, и низкую смертность в потомстве родителей, не связанных кровным родством, в качестве удовлетворительной аппроксимации используется следующая упрощенная формула: S= 1 -A-BF (6.17) 352 6. Популяционная генетика
Вычисления производятся следующим образом: S1 = 1 - A, S2 = 1 - А - FB, S 1 - S2 = BF;
где S1 - число выживающих потомков некровнородственных браков, a S2 - число выживающих потомков в кровнородственных браках. Число летальных эквивалентов определяется как разность между числом мертворожденных и умерших до достижения половой зрелости детей от кровнородственных и некровнородственных браков. Пример. В работе Мёллера, Кроу и Мортона [1827] для предварительной оценки величин А и В использованы некоторые данные по населению Франции (табл. 6.19). Вычисленная для мертворождений и гибели в детстве и юности (до достижения репродуктивного возраста) оценка В находилась в интервале между 1,5 и 2,5; величина А + В была немногим выше. Отношение В/А, которое будет играть важную роль в последующем изложении, варьирует от 15,06 до 24,41. Это означает, что в среднем гамета несет такое число неблагоприятных генов, которое при распределении их между отдельными индивидами и переходе в гомозиготное состояние приведет к гибели до достижения репродуктивного возраста 1,5-2,5 человека. Общий генетический груз составляет 1,5-2,5 летального эквивалента на гамету; 3-5 летальных эквивалентов на зиготу. В этом расчете не учитывались спонтанные аборты и гибель во взрослом возрасте (например, в течение репродуктивного периода). Следовательно, при таком подходе оценка генетического груза получается заниженной. Вероятно, каждый человек гетерозиготен по нескольким мутациям, которые в гомозиготном состоянии оказывают неблагоприятное действие. Авторы осторожно замечают, что разница между кровнородственными и некровнородственными браками может отчасти объясняться причинами небиологического характера. Путем прямого опроса определялся только исход беременности в кровнородственных браках; возможно, что в повышенную смертность детей от кровнородственных браков вносят вклад также социальноэкономические различия между городским и сельским населением. Позднее мы увидим, что это предостережение вполне оправдывается. Оценка выявляемого генетического груза. Следующим пунктом в рассуждениях авторов был вывод, что те же самые гены могут оказывать неблагоприятное воздействие даже в гетерозиготном состоянии, т.е. что их «доминантность» h больше 0. Согласно формуле 6.15, вероятность элиминации данного мутанта в условиях естественной системы скрещивания равна приблизительно zxs, где z = F + q + h (обозначения те же, что и выше). Можно показать, что число выявляемых летальных эквивалентов равно произведению общего числа летальных эквивалентов на гармоническую среднюю величин z для отдельных мутантов. Сведения, необходимые для определения h у человека отсутствовали, поэтому были использованы данные, полученные на дрозофиле. На их основе для 16 аутосомных леталей рассчитаны значения h со средней, равной 0,04. Учитывая, что мутации с более вредным эффектом в природных популяциях должны встречаться реже, и предполагая, что в основном вредное влияние производится гетерозиготами (из-за их более высокой частоты), оценка гармонической средней z для всех вредных генов составляет 0,02. 6. Популяционная генетика 353
При общем числе летальных эквивалентов на гамету, равном 1,5-2,5, это соответствует 3-5% выявляемой летальности на гамету или 6-10% на зиготу. Оценка общей скорости мутирования для неблагоприятных мутаций. Как уже отмечалось, Холдейн (1935) [1472] постулировал существование генетического равновесия между отбором и мутационным процессом. В течение достаточно долгого времени в каждом поколении число вновь возникающих мутаций должно равняться числу вредных аллелей, теряющихся из популяции вследствие их летальности. Отсюда скорость мутирования была также оценена как 0,03 0,05 на гамету на поколение. Авторы предположили, что от 1/2 до 2/3 реального генетического груза невозможно обнаружить путем анализа мертворождений и младенческой смертности (например, невозможно выявить гибель ранних эмбрионов). Принимая это во внимание, была получена оценка общей скорости мутирования, равная 0,06-0,15 на гамету [1827]. Эта величина согласуется с оценкой, приводимой Мёллером в его работе «Наш груз мутаций» [1835]. Однако читатель не должен забывать, что эта оценка основывается на двух допущениях: 1) более высокая частота мертворождений и неонатальная смертность в потомстве кровнородственных браков по сравнению с некровнородственными (проанализированная в упоминавшейся выше работе и ведущая к высокому значению В/А) действительно являются биологическим следствием кровного родства; 2) летальные и вредные гены и в гетерозиготном состоянии уменьшают приспособленность их носителей. Критика выводов, полученных на основе теории генетического груза, касается в основном этих двух предположений. Влияние концепции генетического груза на развитие популяционной генетики человека. Каждый человек гетерозиготен по нескольким генам, которые не только могут привести к генетически детерминированной гибели (особенно в потомстве кровнородственных браков), но неблагоприятны даже в гетерозиготном состоянии. В популяции с высокой скоростью происходит постоянное возникновение новых мутаций, имеющих вредное действие. Можно сказать, что каждый человек менее здоров, чем в том случае, если бы он был свободен от этих мутаций. Концепция генетического груза оказала большое влияние на теоретическое мышление и планирование исследований в области популяционной генетики человека. Отчасти это произошло благодаря присущей данной теории привлекательности, поскольку исследования в этом направлении обещали дать общее представление о проблемах, определяющих будущее нашего вида. Вероятно, большую роль в успехе концепции сыграла также научная репутация разработавшей ее группы ученых: лауреата Нобелевской премии Мёллера, оставившего изучение дрозофилы ради того, чтобы внести свой вклад в спасение человечества; известного популяционного генетика Кроу, чье участие гарантировало надежность подхода, и Мортона, выдающегося молодого ученого с блестящим будущим. Дискуссии и противоречия по поводу концепции генетического груза. Концепция генетического груза широко обсуждалась популяционными генетиками [1809; 1863]. С одной стороны, было установлено, что сравнение потомства кровнородственных и некровнородственных браков может внести вклад в решение вопроса о том, что вносит больший вклад в генетический груз человека - неблагоприятные мутации («мутационный груз») или сбалансированный полиморфизм, обусловленный преимуществом гетерозигот («сегрегационный груз») [1745-1747]. С другой стороны, показано, что в некоторых случаях применение концепции генетического груза приводит к абсурдным выводам [1809]. В настоящее время многие генетики разделяют мнение о том, что пользоваться этой концепцией следует с осторожностью. Современная, несколько более реалистическая версия изложена в работе [1748]. Дата добавления: 2015-12-16 | Просмотры: 1331 | Нарушение авторских прав |
 (6.15)
(6.15)
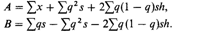
 qs, что соответствует сумме В и генетического компонента А и, следовательно, является промежуточной величиной между В и В + А.
qs, что соответствует сумме В и генетического компонента А и, следовательно, является промежуточной величиной между В и В + А.