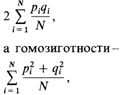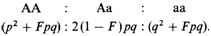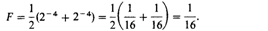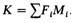|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
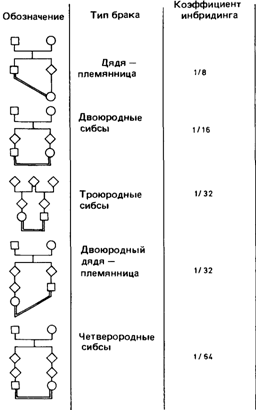
Кровнородственные браки6.3.1.1. Коэффициент инбридинга [103] Все люди – родственники. Родственники определяются как лица, часть генов которых общая по происхождению. Если понимать это определение в буквальном смысле, то родственниками можно считать всех людей. Все мы имеем общих предков. Возможно, ими является одна-единственная пара (см. разд. 7.2.1). Тогда почему же у нас такие разные гены? По той простой причине, что наши общие предки отстоят от нас на тысячи поколений. В течение этого долгого времени произошло множество мутационных событий, которые привели к возникновению генетической изменчивости. Очевидно, считать все человечество родственниками не имеет смысла, так как из этого – пусть даже формально верногопредположения нельзя сделать никаких выводов. Мы измеряем степень кровного родства для того, чтобы исследовать эти мутации и влияние кровного родства на их фенотипическое проявление. Однако при измерении степени кровного родства не следует забывать, что число поколений, которое при этом учитывается, определяется только соображениями практического удобства. Степень родства, которая обычно рассматривается. В большинстве случаев родословные анализируют только по трем поколениям. Это ограничение означает, что рассматриваются только родители, деды и прадеды, а более отдаленные родственники во внимание не принимаются. Таким образом, самые дальние родственники, которые учитываются при оценке кровного родства двух индивидов, это троюродные сибсы. Первоначально это ограничение было введено из чисто практических соображений. Дело в том, что у католиков требуется специальное разрешение на брак между троюродными сибсами или более близкими родственниками и данные по частоте таких браков легко получить из церковных книг, где регистрируются эти разрешения. Типы кровнородственных браков приведены на рис. 6.37. Ограничение оценки кровного родства достаточно узкими рамками оправдано с теоретической точки зрения. При переходе за третье поколение рост коэффициента инбридинга индивида с увеличением числа кровнородственных браков между его (ее) предками происходит очень медленно. Две полезные меры: коэффициент родства и коэффициент инбридинга [130; 1819]. В популяции могут встречаться различные типы кровнородственных браков: между двоюродными и троюродными братьями и сестрами, между дядей и племянницей, а иног- 6. Популяционная генетика 341
да даже между братом и сестрой или отцом и дочерью. Конечно, частоты всех этих типов браков можно оценить: такие данные представляют интерес с социологической точки зрения. Однако для генетика интересен только один аспект данного вопроса: какова степень родства между супругами, какая часть генов у них общая? Если мы хотим сравнить особей по степени инбридинга или описать популяцию по среднему уровню инбридинга составляющих ее особей, нам необходима величина, измеряющая эту общую часть генов. Введение соответствующего параметра упрощает нашу задачу точно так же, как введение понятия генных частот упрощает описание популяции в терминах генотипов. Предложено несколько мер уровня инбридинга; выбор между ними в большой степени условен. Наиболее удобной оценкой является «коэффициент инбридинга» Райта [1885]. Он тесно связан с «коэффициентом родства» Малеко [130]. Эти коэффициенты определяются следующим образом: а) коэффициент родства ФАВ двух индивидов А и В – это вероятность того, что случайно выбранный ген, принадлежащий А, идентичен гену того же локуса у В; б) коэффициент инбридинга индивида равен коэффициенту родства его отца и матери. Разница между этими коэффициентами состоит в том, что коэффициент родства определяется для двух индивидов, которые могут иметь общих предков. Коэффициент инбридинга определяется для одного индивида и измеряет степень связи между его родителями и, следовательно, сходство между двумя генами каждого локуса, полученными от родителей. Коэффициент инбридинга - это вероятность того, что два аллеля в данном локусе идентичны по происхождению. Коэффициент инбридинга и закон Харди — Вайнберга. Рассмотрим два аллеля А и а аутосомного гена; пусть частота этих аллелей равна р и q. В случайно скрещивающейся популяции частоты трех генотипов равны p2:2pq:q2. Если генотип содержит N пар аллелей, а их частоты в популяции равны р i, и q i (i = 1, 2,..., N), степень гетерозиготности при случайном скрещивании будет равна
причем их сумма равна 1. Степень гетерозиготности отражает долю аутосомных генов, по которым в среднем гетерозиготен индивид. В случае одного гена она определяет вероятность того, что данный индивид гетерозиготен по этому гену. Рассмотрим пару аллелей А и а в слу- 342 6. Популяционная генетика
чае кровнородственного брака (рис. 6.38). Пусть ооцит несет аллель а. Если скрещивание случайно, то вероятность того, что этот ооцит будет оплодотворен сперматозоидами, несущими аллели а и А, равна р и q соответственно. Если супруги являются родственниками, то часть их генов имеет общее происхождение, поэтому р увеличивается до (р + Fq), a q уменьшается до (q — Fq). Аналогичные рассуждения справедливы для ооцитов, несущих аллель А. Величину, соответствующую F в предыдущем изложении, здесь можно обозначить как F'. Если наследование аутосомное, распределение генов А и а будет идентичным у обоих родителей. Поэтому pq (1— F) = = qp( 1 — F') и, следовательно, F = F'. Можно показать, что F равен коэффициенту инбридинга, как он был определен выше. Это означает, что частоты генотипов детей, коэффициент инбридинга которых равен F, не соответствуют соотношению Харди—Вайнберга и равны
Степень гетерозиготности ребенка уменьшается в среднем в F раз. Говоря другими словами, F- вероятность того, что две гомологичные хромосомы несут в случайно выбранном локусе два аллеля, происходящие от одного и того же предкового гена. Вычисление коэффициента инбридинга F. В большинстве реальных ситуаций, возникающих в генетике человека, вычисление Ф или F необязательно, поскольку степень кровного родства в популяциях человека обычно известна. Иногда коэффициент инбридинга необходимо вычислить для какой-либо конкретной родословной. Совершенно иное положение в селекции животных, где между скрещиваемыми особями встречаются очень сложные родственные связи. Для определения коэффициента инбридинга Райт разработал метод коэффициентов путей [124; 961]. Для этого строятся родословные скрещивающихся особей и отмечаются все их общие предки. Затем выбирается один из ближайших общих предков и родительские особи соединяются всеми возможными путями, которые: а) ведут к этому общему предку; б) состоят из «шагов» (один шаг определяется как связь между особью и одним из ее родителей); в) не ведут к одной особи более одного раза. Все остальные общие предки анализируются тем же способом. Для каждого пути подсчитывают число шагов. Для одного предка может существовать х путей, состоящих из т 1 ,..., тх шагов каждый; для t общих предков существует
(6.12) 1) Эту формулу легко понять с помощью следующего простого рассуждения. 1/2 генов ребенка общая с каждым из его родителей, ¼ - с дедом и бабкой, 1/8 - с прадедом и прабабкой и т. д. Если от матери к какому-нибудь предку ведет путь из а шагов, она имеет с этим предком (1/2) a = 2–a общих генов. Если от отца к этому предку ведет путь из b шагов, он имеет с этим предком 2–b общих генов. Это означает, что отец и мать имеют 2–a х 2–b — 2–т общих генов (т = а + b). Это число, деленное на 2, дает вероятность того, что ген, случайно выбранный у матери, идентичен по происхождению гену, случайно выбранному у отца 2). Примеры. На рис. 6.39 показан брак между двоюродными сибсами. Пути к общему деду и общей бабке у этой пары состоят каждый из 4 шагов. 1) Если один из общих предков происходит от кровнородственного брака, необходимо внести поправку с учетом коэффициента инбридинга. 2) Более строгий вывод и другие методы вычисления F см. в [124; 103; 108]. 6. Популяционная генетика 343 Подстановка в уравнение (6.12) дает
На рис. 6.40 показан брак отца и дочери. Здесь только один путь, состоящий из одного шага:
Третий пример - брак между братом и сестрой (рис. 6.41):
Как упоминалось выше, для человека эти вычисления условно ограничены тремя поколениями. Это условие разумно с количественной точки зрения: например, если общий предок отстоит от данной пары на 5 поколений, соответствующий путь имеет 5 шагов; следовательно, его вклад в F равен только 1/2·2–10 = 1/2048.
Коэффициент инбридинга популяции. Во многих случаях нас интересует величина, измеряющая среднюю степень кровного родства в популяции и учитывающая все типы имеющихся в ней кровнородственных браков. Если ограничить анализ последними тремя поколениями, то в результате мы получим коэффициент явного кровного родства К:
Суммирование производится по различным типам кровнородственных браков; Fi и М i - коэффициент инбридинга и относительная частота i -того типа кровнородственного брака. К зачастую называют просто средним F или F популяции. При сравнении по этому параметру разных популяций необходимо учитывать, что условие, касающееся анализа только трех ближайших поколений, соблюдается не всегда. Кроме того, почти во всех популяционных исследованиях определяют не коэффициенты инбридинга всех особей, а коэффициенты родства всех пар. Коэффициент родства дает несмещенную оценку коэффициента инбридинга только в том случае, когда инбридинг не влияет на репродукцию. 344 6. Популяционная генетика Дата добавления: 2015-12-16 | Просмотры: 1542 | Нарушение авторских прав |