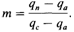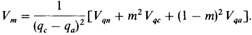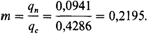|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Дифференциация субпопуляций: генетическое расстояниеРеальная структура скрещивания популяций человека. Обычное предположение о том, что в популяциях человека преобладает случайное скрещивание - это абстракция. Выбор брачного партнера вовсе не случаен. Он зависит от расстояния между местами рождения вступающих в брак, от ограничений, обусловленных религией, расовой, языковой и социальной принадлежностью и т.д. В прошлом неслучайные браки заключались гораздо чаще, чем сейчас. Особенно ярко выраженным был неслучайный выбор партнеров в течение тысяч поколений доисторического времени, когда наши предки жили как охотники и собиратели, объединяясь в маленькие группы. Изучение современных племен дает возможность получить представление о структуре популяций того времени. Небольшие популяции, находящиеся в относительной изоляции друг от друга, постепенно станут различаться генетически - либо в результате давления отбора различной интенсивности, либо благодаря разным способам адаптации к одному и тому же селективному фактору, либо просто из-за случайной флуктуации генных частот (разд. 6 4). Конечная цель популяционной генетики - анализ причин таких популяционных различий. Однако в большинстве случаев эта цель оказывается недостижимой. В отсутствие более определенной информации во многих случаях удобно начать с предположения, что генетическое сходство между популяциями обусловлено их общим происхож- 364 6. Популяционная генетика дением. Отсюда, чем более сходны две популяции, тем ближе они генетически. Это означает, что если популяции сходны, то они разделились недавно, если же их сходство не очень велико, можно сделать вывод, что с момента обособления прошло более длительное время. Следовательно, на основании генетического состава популяций, наблюдаемого в настоящее время, можно сделать заключение об истории этих популяций. В принципе этот метод лежит в основе любой классификации (например, расы) у людей. В разд. 6.2.1.7 рассмотрен хорошо известный пример такого рода: распространение аллеля HbF среди населения, принадлежащего южноазиатской языковой группе. Здесь мутация HbβE достигла высокой частоты благодаря селективному преимуществу ее носителей, их устойчивости к малярии. Эта мутация «мигрировала» вместе с субпопуляциями данной группы в различные области страны, и теперь высокая частота НbЕ свидетельствует не только о продолжающемся отборе под влиянием малярии, но также и о происхождении от определенной популяционной группы. Таким образом, аллель HbβE является свидетелем истории популяции. История популяции или отбор? На этом примере можно судить и о том, насколько неоднозначным может быть распределение генетического признака. Аллель HbβE обычно реже встречается у населения равнин великих рек Юго-Восточной Азии по сравнению с населением холмистых областей, где малярия более распространена. Однако эта дифференциация не имеет никакого отношения к разделению популяций, произошедшему в отдаленные исторические периоды; она возникла в относительно недавнее время просто в результате менее интенсивного давления отбора, обусловленного малярией. Генетическое сходство между популяциями тоже можно интерпретировать по-разному. Оно может отражать (но не обязательно отражает) общую историю этих популяций. С другой стороны, такое сходство может свидетельствовать о параллельном изменении генофонда популяций под влиянием сходного давления отбора. В рассмотренном примере, касающемся населения Юго-Восточной Азии, в качестве показателя сходства между популяциями использовался только один ген. Можно предположить, что чем большее число различных генов вовлечено в такой анализ, тем менее вероятность того, что все они находятся под одинаковым давлением отбора. Следовательно, сходство популяций по большому числу генов убедительно свидетельствует в пользу общего происхождения этих популяций. Действительно, при анализе сходства и различий между популяциями обычно стараются использовать как можно больше наследственных признаков и применяют методы многомерной статистики. Тем не менее, рассматривая выводы таких исследований, необходимо всегда иметь в виду, что обычно о селективных силах, оказывающих влияние на популяцию, ничего не известно. Методы определения генетического расстояния. Методы определения генетического расстояния между популяциями обсуждаются в нескольких прекрасных обзорах [36; 103; 1893]. Для непрерывно распределенных признаков, например антропологических измерений, обычно используют обобщенное расстояние Махалонобиса, его упрощенный вариант - индекс Пенроуза D2. Для альтернативных признаков, таких как полиморфные признаки с простым типом наследования, как правило, применяются угловая и хордовая меры Кавалли-Сфорца и Эдвардса (1964) [1735]. Поток генов Кроме отбора (рассмотренного выше) и случайных флуктуации генных частот (которые будут обсуждаться позднее), на генофонд популяции оказывает большое влияние также поток генов. Для обозначения переноса генов из одной популяции в другую часто применяется термин «миграция». Влияние миграции на генные частоты [124]. Действие миграции на частоты генов будет рассмотрено на несколько упрощенной модели. Большая популяция может подразделяться на субпопуляции меньшего размера. Предположим, что средняя частота гена равна
Δ q пропорционально как отклонению частоты гена в субпопуляции q от средней частоты в популяции в целом ( 6. Популяционная генетика 365 ние отбора в субпопуляциях) различия между субпопуляциями сгладятся и все они будут иметь одну и ту же частоту гена Миграция и отбор. Если субпопуляции подвержены давлению отбора различной интенсивности, этот отбор может противостоять процессу выравнивания генных частот. Здесь возможны три ситуации 1. 1. Если скорость миграции и интенсивность отбора имеют один порядок величины, частоты генов в субпопуляциях могут сильно отличаться друг от друга. 2. Если интенсивность отбора намного превышает скорость миграции в субпопуляциях, частоты генов в субпопуляциях будут определяться в основном отбором и «разбавляющее» влияние миграции окажется очень слабым. 3. Напротив, если доля «иммигрантов» намного выше интенсивности отбора, действие миграции «перевесит» действие отбора. В любом случае возможно установление стабильного равновесия между отбором, с одной стороны, и миграцией, с другой. Такая ситуация несколько сходна с равновесием между отбором и мутационным процессом (разд. 5.2). Измерение притока генов в субпопуляцию. Часто производится оценка доли генов, которую субпопуляция получила извне путем миграции. Пусть qa – частота гена в «чистой» предковой популяции, a qn – частота того же гена в той же популяции в настоящее время; предполагается, что в эту популяцию произошел приток генов извне. Пусть частота гена в популяции-«доноре» равна qc. Тогда доля т генов в исследуемой популяции в настоящее время, пришедших из популяции-«донора», составляет qп = тq с+ (1 - m)qa и, следовательно,
Вариансу т можно определить следующим образом:
Оценка притока генов в субпопуляцию. В последнее время всеобщее внимание привлекает вопрос о том, какова доля генов белого населения (и других расовых групп) у негров США. Хотя в принципе оценить это просто, решение данной задачи требует выполнения определенных условий [1857]: а) должны быть известны точный этнический состав предковой популяции и частоты генов, используемых для оценки; б) частота анализируемого гена (генов) не должна систематически изменяться в течение времени, прошедшего от «предкового» поколения до настоящего времени. Систематическое изменение генных частот может быть вызвано естественным отбором. Например, в негритянском населении Америки распространен ген серповидноклеточности. Известно, что этот ген широко распространен в Африке благодаря отбору в связи с малярией (разд. 6.2.1.6); в Северной Америке такого отбора нет. С другой стороны, в США ген серповидноклеточности должен испытывать направленное против него давление отбора, возникающего в результате выщепления гомозиготных индивидов, страдающих серповидноклеточной анемией. Таким образом, оценка, основанная на частоте этого гена, приведет к завышению примеси генов белой расы. Однако можно рассуждать и иначе. Если оценка примеси генов производится на основе одного или (в идеальном случае) многих генов, удовлетворяющих этим ус-
1 Математический анализ этих вопросов см. в работах [103; 124]. 366 6. Популяционная генетика ловиям, разница между этой оценкой, и оценкой, произведенной на основе гена, испытывающего давление отбора, может быть использована для демонстрации наличия отбора и измерения его интенсивности. Оценка притока генов белой расы в популяцию американских негров. Американские негры произошли от рабов, завезенных в США из Западной Африки (Нигерии, Сенегала, Гамбии, Кот-Дивуара, Либерии и др.). В этих предковых популяциях частоты большинства генетических маркеров обнаруживают ярко выраженную изменчивость. То же самое справедливо и для белого населения Америки, которое произошло от иммигрантов из различных областей Северной, Западной, Центральной и Южной Европы. Возможно, что гены, внесенные в генофонд американских негров, не являются несмещенной и случайной выборкой генов всего белого населения США. Некоторые группы белого населения могли принимать более активное участие в аутбридинге по сравнению с другими группами. Все же тщательное выявление возможных отклонений способствует определению правильного порядка величины притока генов [1857]. Считается, что дифференциальный отбор почти или совсем не оказывает влияния на оценки притока генов, основанные на системах групп крови и сывороточных белков (Rh, Duffy, группы Gm). Для различных негритянских субпопуляций эти оценки изменяются от т = 0,04 до т = 0,30. Величины т для населения южных сельских районов, как правило, ниже, чем для крупных городов типа Балтимора или Нью-Йорка, для которых они обычно превышают 0,2. Рассмотрим метод оценки притока генов на следующем примере. Частота аллеля Fya групп крови системы Duffy составляет у американского белого населения qc = 0,43. В популяциях Западной Африки его частота qa в настоящее время ниже 0,03, а в большинстве групп населения Африки Fya вообще отсутствует. Можно предположить, что в то время, когда происходил вывоз рабов из Африки, частота аллеля Fya также была очень низкой. В современном негритянском населении Окленда, штат Калифорния (и = 3,146), частота Fya q n = = 0,0941 + 0,0038, соответствующая частота в белом населении (и = 5 046) qc = = 0,4286 + 0,0058; qa (частота гена в африканской популяции) принимается равной 0. Пользуясь формулой, приведенной выше, получим следующую оценку притока генов:
Если принять qn за 0,02, эта оценка составит 0,181. Следовательно, приток генов белого населения, определенный на основе частот групп крови Duffy, составляет 18-22% генофонда негритянского населения Окленда (Калифорния). Оценка притока генов на основе системы АВО для той же популяции приводит к сходному результату (т = 0,20). Доказательство наличия отбора. Как отмечалось, оценки притока генов, полученные для генов, испытывающих в африканских популяциях давление отбора, можно использовать для определения того, изменились ли интенсивность и направление отбора в новом месте обитания африканских негров. В нескольких работах были получены более высокие оценки притока генов белых для трех генетических маркеров: гена серповидноклеточности (Hb|3S), аллеля африканского варианта G6PD (GDA‑) и аллеля гаптоглобина Нр 1. Как отмечалось в разд. 6.2, на аллели HbβS и GdA‑ в Африке действует отбор, связанный с тропической малярией; гаптоглобин является белком, участвующим в переносе гемоглобина. Определенные величины притока генов для этих аллелей были достоверно выше, чем соответствующие оценки, полученные на основе частот систем групп крови Duffy и АВО; они варьировали приблизительно от 0,49 (GdA‑, Сиэтл, северо-запад США) до 0,17 (GDA‑, Мемфис, юг США). Эти результаты указывают на то, что в Соединенных Штатах - стране, где малярии нет,против этих генов идет отбор. Из-за отсутствия необходимых данных интенсивность этого отбора не может быть точно определена. Для доказательства существования отбора в отношении генов, для которых 6. Популяционная генетика 367 оценка притока генов таким способом невозможна, необходимо использовать другие подходы. Дата добавления: 2015-12-16 | Просмотры: 1166 | Нарушение авторских прав |
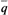 ; каждая субпопуляция в каждом поколении обменивается со случайной выборкой из всей популяции долей своих генов, равной т. Пусть генная частота в первом поколении в рассматриваемой популяции равна q. Тогда в следующем поколении частота гена в этой субпопуляции будет равна
; каждая субпопуляция в каждом поколении обменивается со случайной выборкой из всей популяции долей своих генов, равной т. Пусть генная частота в первом поколении в рассматриваемой популяции равна q. Тогда в следующем поколении частота гена в этой субпопуляции будет равна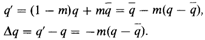
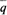 ), так и т. С течением времени в отсутствие других факторов (таких как дифференциальное давле-
), так и т. С течением времени в отсутствие других факторов (таких как дифференциальное давле-