|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Отбор, приводящий к нестабильному равновесию
Отбор против гетерозигот. В популяции может установиться стабильное равновесие, если отбор идет в пользу гетерозигот и против гомозигот. Однако отбор может действовать и в пользу гомозигот против гетерозигот. Пусть приспособленности генотипов АА, Аа и аа равны 1, 1 — s и 1 соответственно. Изменение генных частот в этом случае показано в табл. 6.8. Отсюда
если s мало. На рис. 6.11 показано изменение Δ q в зависимости от q. Если чие генетического равновесия, однако в данном случае равновесие неустойчивое. Если q больше1/2, Δ q положительно. Частота гена стремится к 1. С другой стороны, если q меньше1/2, Δ q становится отрицательным и q стремится к 0. Нестабильное равновесие само по себе неспособно поддержи- 6. Популяционная генетика 303
вать в популяции полиморфизм; даже в условном примере, где две гомозиготные по двум разным аллелям популяции смешиваются в равных пропорциях, небольшие сдвиги генных частот, происходящие в результате случайных флуктуации, скоро приведут к нарушению состояния равновесия и частота гена начнет сдвигаться к 1 или 0. Тем не менее такая маловероятная ситуация, возможно, имеет место. Для человека известно два случая, когда единственный селективный фактор, который пока установлен, - это отбор против гетерозигот. В одном из этих случаев, системе Rh (11170), аллельные частоты соответствуют нашему определению полиморфизма. В случае перицентрических инверсий гетерозиготы имеют полиморфные частоты и иногда встречаются гомозиготы. Перицентрические инверсии. У гетерозигот по перицентрической инверсии спаривание гомологичных хромосом во время мейоза может произойти неправильно. Образующиеся в результате несбалансированные гаметы элиминируются перед оплодотворением или образуют нежизнеспособные зиготы. С другой стороны, у гомозигот по перицентрическим инверсиям нарушений мейоза не ожидается, у них спаривание гомологов должно проходить нормально. Действительно, в потомстве родителей, имеющих перицентрические инверсии, наблюдается высокий уровень репродуктивной потери [401а]; соответствующие популяционно-генетические исследования не проводились. Однако такие исследования необходимы, поскольку перицентрические инверсии играют важную роль в эволюции человека: по-видимому, они обеспечивают мощный механизм репродуктивной изоляции. Перицентрические инверсии не влияют на состояние здоровья гетерозиготных носителей, однако ухудшают их репродуктивную способность. Это классический (и, возможно, лучший) пример влияния плодовитости на приспособленность при отсутствии изменения жизнеспособности. Вероятно, этим и определяется значение перицентрических инверсий для эволюции и в особенности для видообразования (разд. 7.2.1). Отбор против гетерозигот по Rh-фактору. Неустойчивое равновесие, возникающее при отборе против гетерозигот, было впервые открыто Холдейном (1942) [1777] для конкретного случая серологической несовместимости матери и ребенка по Rh-фактору. Этот случай несколько сложнее, чем рассмотренная выше ситуация с перицентрическими инверсиями. Угроза эритробластоза и, следовательно, отбор против гетерозигот возникают, если Rh-отрицательная мать вынашивает Rh-положительный плод. Локус Rh имеет сложную структуру. Однако для понимания принципа отбора по этому локусу достаточно рассмотреть только аллели D (положительный) и d (отрицательный), что сводит задачу к простой диаллельной системе. Дети с эритробластозом рождаются у родителей dd♀ x DD♂ или dd♀ х Dd♂. В табл. 6.9 приве-
304 6. Популяционная генетика дены типы брачных пар; строчки, соответствующие детям, против которых действует отбор и для которых велик риск возникновения эритробластоза, выделены цветом. Учитывая все типы браков, получаем следующую формулу для изменения частоты аллеля D:
Здесь s 1отражает отбор против детей, матери которых имеют генотип dd, a отцы – DD; s2 – отбор против гетерозиготных детей матерей dd и отцов Dd. Поскольку риск иммунизации возрастает с числом детей генотипа Dd, он ниже, если отец имеет генотип Dd (ведь в таком браке в среднем только каждый второй ребенок вызывает у матери иммунизацию), тогда как у отца с генотипом DD каждый ребенок гетерозиготен и может привести к иммунизации матери; следовательно; s2 < s 1. Легко показать, что Δ р = р' — р только в том случае, когда р =1/2. Это означает, что уравнение (6.8) имеет ту же точку равновесия, что и уравнение (6.7). Здесь равновесие также нестабильно (рис. 6.12). В современных популяциях Западной Европы аллель d встречается с частотой q = 0,35. Следовательно, его частота должна уменьшаться при отсутствии других селективных механизмов, противодействующих этой тенденции. С какой скоростью будет происходить это уменьшение частоты? На рис. 6.13 показано Δ р для нескольких поколений, полученное при двух предположениях о начальных частотах р для D и q для d, и коэффициентах отбора s1 и s2, изменяющихся в пределах, действительно наблюдаемых в популяциях человека. Перед введением профилактической терапии сывороткой анти-D у женщин из группы риска число пораженных детей зависело от среднего числа беременностей [1740]. Правдоподобной оценкой является 5% детей, пораженных эритробластозом, от всех детей Dd матерей dd. В прежнее время, когда женщины имели больше беременностей, эта цифра, вероятно, была несколько выше. Из рис. 6.13 видно, что изменение генных частот в этих условиях происходит очень медленно из-за того, что не все гетерозиготы подвержены отбору (табл. 6.9). Отбор против гетерозигот может нарушаться другими, пока неизвестными формами отбора, даже если их интенсивность очень низка. Возможно, что время, в течение которого в популяциях происходило случайное скрещивание, недостаточно для проявления действия отбора против гетерозигот (имеется в виду время, прошедшее с
6. Популяционная генетика 305
того периода, когда случайные флуктуации в популяциях человека, состоящих из небольших сравнительно изолированных субпопуляций, создавали огромные межпопуляционные различия генных частот - разд. 6.4.1). Исчерпывающее объяснение полиморфизма по Rh-фактору с точки зрения популяционной генетики остается загадкой ввиду высокой частоты аллелей системы Rh в очень многих популяциях. Предполагается, что в отборе по этой системе принимают участие другие пока неизвестные «силы». Система групп крови АВО. Серологическая несовместимость матери и плода характерна также для системы групп крови АВО. В отличие от Rh-иммунизации здесь может быть затронут даже первый ребенок [211]. Эритробластоз в этом случае, как правило, выражен слабее, чем при Rh-несовместимости. Однако АВО-несовместимость может также привести к увеличению числа спонтанных абортов, хотя данные об этом противоречивы [211]. Очевидно, отбор идет в основном против детей АО и ВО матерей 0. Соответствующие рекуррентные уравнения можно получить аналогично приведенным выше для двухаллельного случая системы Rh. Разница состоит только в том, что в системе АВО отбор против гетерозиготных детей гомозиготных и гетерозиготных отцов одинаков, поскольку в отличие от Rh-иммунизации первый ребенок с несовместимостью может быть уже пораженным. Изменение частоты аллеля А описывается следующей формулой:
Совершенно аналогичная формула может быть получена для Aq (изменение частоты аллеля В). Формула для Аг, изменения частоты аллеля 0, несколько отлична:
(6.10) Если r = 0,5, Δ р = Δ q = Δ r = 0 независимо от соотношения между р и q. Это равновесие нестабильно по отношению к r и нейтрально по отношению к p и q. На рис. 6.14 показана скорость, с которой r приближается к 1, a p и q – к 0. Предполагается, что другие селективные механизмы не действуют; начальные аллельные частоты приняты равными соответствующим частотам в населении Западной Европы (А: p = 0,3; В; q = 0,16; 0: r = 0,6). Коэффициенты отбора получены на основе эмпирических данных. Изменение генных частот происходит гораздо быстрее, чем в случае Rh (рис. 6.13). Дата добавления: 2015-12-16 | Просмотры: 696 | Нарушение авторских прав |
 (6.7)
(6.7) Δ q = 0. Это означает нали-
Δ q = 0. Это означает нали-

 , и положительно, если q >
, и положительно, если q > 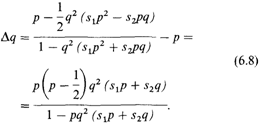


 (6.9)
(6.9)