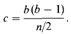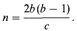|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Инбридинг, размер изолята и наследственные заболеванияЧастота детей с рецессивными и полигенными заболеваниями в кровнородственных и неродственных браках. Пусть аллель, который в гомозиготном состоянии приводит к возникновению рецессивного заболевания, встречается в популяции с частотой q. Тогда частота соответствующего фенотипа в случайно скрещивающейся популяции будет равна q2; если коэффициент инбридинга особей в популяции равен F, эта частота составит q2 + Fpq. С уменьшением q увеличивается отношение Fpq/q2: чем ниже частота гена (генотипа), тем выше частота кровнородственных браков среди родителей пораженных гомозигот. Этот вывод справедлив не только для рецессивных заболеваний, но и для полигенных признаков (разд. 3.6). У особей с коэффициентом инбридинга F дисперсия нормально распределенной подверженности заболеванию с наследуемостью h2 = 1 равна VF=V 0 ( 1 + F), где V 0 — дисперсия в неинбредной популяции. Однако с увеличением дисперсии относительное число особей со значением признака, превышающим пороговое, также увеличится. Следовательно, риск заболевания мультифакториальной болезнью с пороговым эффектом несколько выше для детей от кровнородственных браков, чем для детей, родители которых не являются родственниками (рис. 3.63). В случае рецессивных генов ситуация противоположная. Если в популяции обнаружено большее число кровнородственных браков, чем ожидалось на основании частоты встречаемости в ней определенного рецессивного заболевания, возможно, что это заболевание детерминируется не одним, а несколькими рецессивными генами, имеющими более низкие частоты, т. е. существует генетическая гетерогенность. Fxли предположить, что частоты этих генов одинаковы, можно даже определить их число. Однако на практике такой подход почти всегда оказывается бесполезным по следующим причинам. 1. Уменьшение инбридинга в современных популяциях привело к резкому снижению числа гомозигот по рецессивным заболеваниям. 2. Уменьшение инбридинга характерно главным образом для больших городов и территорий с большой плотностью населения. В отдаленных сельских областях, где кровнородственные браки встречаются чаще, рецессивные заболевания проявляются. 3. Высокое значение отношения кровнородственных браков к неродственным может объясняться просто гетерогенностью популяции даже при наличии только одного рецессивного гена. Эти вопросы будут обсуждаться в связи с генетической природой глухонемоты (приложение 3). Определение величины изолята по частоте кровнородственных браков. Если браки заключаются случайным образом, а размер популяции бесконечно большой, вероятность кровнородственного брака равна нулю. Это утверждение можно сформулировать противоположным образом. Если браки внутри изолята происходят случайно, какова должна быть его величина, чтобы соответствовать реально наблюдаемой в этом изоляте частоте кровнородственных браков? Эта идея лежит в основе определения размера изолята [1750]. Пусть n -число индивидов в изоляте, n /2число индивидов каждого пола, способных вступить в брак, а b - среднее число детей на брак. Тогда среднее число двоюродных сибсов противоположного пола для каждого индивида будет равно b (b — 1). Если вероятность их вступления в брак такая же, как и у других членов популяции, то доля кровнородственных браков составит
Решая это уравнение, для п получаем
Такие же вычисления можно произвести для других степеней родства. Они широко применялись разными авторами для определения структуры скрещиваний в популяциях человека. В действительности кровнородственные браки происходят не случайно: на них влияет множество различных социальных и социально-пси- 6, Популяционная генетика 345
хологических факторов. Слабость этого метода можно легко продемонстрировать, показав, что оценки размера изолята, основанные на различных типах кровнородственных браков, дают совершенно разные результаты для одной и той же популяции (табл. 6.17). Коэффициент инбридинга F в различных популяциях. В табл. 6.18 приведены частоты кровнородственных браков в различных популяциях. В этой таблице приведены браки между двоюродными сибсами (1–С) и значения F, вычисленные на основе имеющихся данных 1). Сведения получены при анализе родословных пар, заключивших кровнородственный брак. В зависимости от метода оценки значение коэффициента инбридинга F может быть в той или иной степени занижено по следующим причинам. 1. Для стран с католическим населением данные обычно основаны на записях разрешения на брак, которые имеются в церковных книгах. Однако, вероятно, не все католики, собирающиеся вступить в брак с близким родственником, обращаются в церковь за разрешением; священники, особенно в городах, могут недостаточно хорошо знать своих прихожан, чтобы определить родственные связи между ними. 2. В некоторых работах используются опросы семей, но надо учитывать, что кровное родство между супругами часто скрывается. Пример: при изучении населения острова Хосодзима (Япония) семь членов совета острова из общего числа браков 45 выделили 19 кровнородственных. Проверка регистрационных записей увеличила это число до 25, в результате тщательного анализа родословных число браков между родственниками возросло до 29 [1794]. Почти во всех странах Европы и в США коэффициенты инбридинга очень низкие; высокие коэффициенты инбридинга характерны для небольших поселений, а также для религиозных, географических и этнических изолятов. В Южной Америке, которая изучена очень хорошо, средний коэффициент инбридинга, вероятно, в два или три раза выше, чем в Европе [1763; 1764]. Высокие значения F показаны также и для Японии. Наиболее высокие значения получены для населения некоторых областей Южной Индии (особенно для штата Андра Прадеш), нубийских племен Египта и племени фульбе Гвинеи. Падение частоты кровнородственных браков в развитых странах. В странах Западной Европы с высоко развитой промышленностью с начала XX в. наблюдалось уменьшение частоты кровнородственных браков. Эта тенденция зародилась в индустриальных областях и больших городах и сейчас распространяется в сельскохозяйственные районы. Особенно подробно в этом отношении изучена Франция. В 1926-1930 гг. средний коэффициент инбридинга (точнее,
1) Для Израиля приведены только частоты браков двоюродный брат—двоюродная сестра, дядя—племянница и тетя—племянник, поэтому значение коэффициента инбридинга может быть заниженным.
348 6. Популяционная генетика
коэффициент явного кровного родства, см. выше) был равен 86,1 х 10–5; в 19561958 гг. он упал до 23 х 10–5. Уменьшение коэффициента инбридинга обычно объясняют повышением мобильности населения промышленно развитых стран и расширением возможности выбора брачного партнера. Такое объяснение подтверждается исследованиями, в которых показано, что расстояние между местами рождения супругов возрастает во времени. Это явление обычно называется «разрушением» изолятов. В последнее время тенденция к уменьшению числа родственных браков усилилась из-за уменьшения числа детей в семьях и, следовательно, уменьшения числа двоюродных сибсов. Исходя из этих и других соображений, возникает вопрос о том, в какой степени кровнородственные браки представляют собой репрезентативную в других отношениях выборку из всех браков. Этот вопрос имеет значение при использовании кровнородственных браков для оценки генетического груза (разд. 6.3.2), обусловленного летальными и неблагоприятными в гомозиготном состоянии генами. Влияние социальных и психологических факторов на частоту кровнородственных браков. В промышленно развитых районах Франции, где частота кровнородственных браков ниже, качество акушерской и педиатрической помощи обычно выше. Исследования, проведенные в Германии, показали, что люди, вступившие в брак с близким родственником, в психологическом отношении отличаются от среднепопуляционного уровня [1896]. Например, мужья из кровнородственных браков с большим трудом, нежели другие мужчины, устанавливали межличностные контакты, именно поэтому они и выбрали в качестве жены родственницу. В некоторых регионах отношение к кровнородственным бракам положительное, например в Южной Индии наиболее предпочтительным с социальной точки зрения считается брак между племянницей и ее дядей по материнской линии. До самого недавнего времени в Индии и Японии выбор жениха и невесты осуществлялся их семьями. Определяющую роль при этом играли экономические мотивы, а также степень знакомства с семьей жениха (невесты). В этом случае психологические факторы, подобные упомянутым выше в связи с исследованием популяции Германии, вероятно, играют небольшую роль. По этой причине, а также по ряду других соображений можно сделать вывод, что в Японии кровнородственные браки являются более репрезентативной выборкой из всех браков по сравнению с Европой. Однако некоторые смещения наблюдаются и здесь. Например, изучение кровнородственных браков в населении японского острова Хирадо [1877; 1878; 1879] показало повышенную частоту браков между старшими братьями и их двоюродными сестрами. Кроме того, именно старшие братья наследовали участок земли, принадлежащий семье, поэтому они чаще оставались в родной деревне, тогда как младшие братья меняли место жительства. При определении влияния кровного родства родителей на состояние здоровья потомства такие отклонения должны учитываться. Влияние уменьшения частоты кровнородственных браков на частоту рецессивных заболеваний. Предположим, что в популяции со средним коэффициентом инбридинга F при частоте гена, равной q, скорость мутирования μ и коэффициент отбора s находятся в состоянии равновесия. Пусть в течение короткого промежутка времени, равного, скажем, одному поколению, коэффициент инбридинга уменьшился от F lдо F2. Число гомозигот соответственно упадет с q2 + F 1 pq до q2 + F 2 pq. Это изменение частот нарушит генетическое равновесие. Поскольку гомозигот стало меньше, давление отбора теперь недостаточно для элиминации возникающих в результате мутирования генов. Например, для населения Европы F 1варьирует в пределах 0,003-0,005. Предположим, что скорость мутирования равна 10–5, а коэффициент отбора (s) против гомозигот – 0,5. Равновесная частота q = 2,6-3,2 х 10–3. В этом случае полное прекращение инбридинга приводит к падению частоты гомозигот, показанному на рис. 6.42. Для того чтобы пройти половину пути к достижению новой равновесной час- 6. Популяционная генетика 349
тоты, популяции понадобится 175-185 поколений, или около 4500 лет. Уменьшение инбридинга является одной из причин того, что в современном обществе рецессивные наследственные болезни встречаются и будут встречаться еще в течение долгого времени с такой необыкновенно низкой частотой. Вторая причина, связанная с первой, заключается в том, что в результате стохастических процессов в популяциях малого размера (разд. 6.4) частоты генов, определяющих рецессивные заболевания, различны в разных группах населения. При увеличении числа браков между представителями этих групп генные частоты в них выравниваются, а высокие частоты генов и соответственно высокие частоты гомозигот исчезают. Дата добавления: 2015-12-16 | Просмотры: 820 | Нарушение авторских прав |