|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
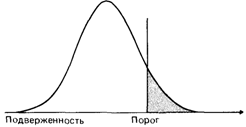
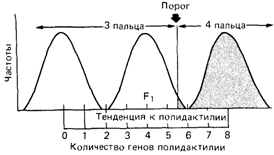
Описание модели: эксперименты на животныхВ предшествующем разделе генетический анализ количественного признака на биометрическом уровне обсуждался в отношении нормальных признаков с унимодальным и почти нормальным распределением в популяции. Было показано, что простая модель аддитивною полигенного наследования удовлетворяет этим свойствам, и тем самым корреляции родитель-ребенок и сибс-сибс можно использовать для оценки наследуемости. Однако для многих болезней и врожденных пороков развития наблюдаются четкие альтернативные распределения: индивид либо страдает данным заболеванием, либо нет. Между тем ни семейные исследования, ни изучение хромосом не смогли выявить какой-либо простой тип наследования или наличие хромосомной аномалии. Единственное, что следует из семейных данных,это возрастание эмпирического риска для близких родственников оказаться пораженными тем же заболеванием (семейное накопление). Патофизиологические исследования позволяют предполагать сложный комплекс причин. В некоторых случаях очевидно влияние различных дополнительных биологических факторов. Осложнения привносят и средовые факторы: неправильное питание, инфекции и неизвестные агенты. Когда все эти генетические и средовые факторы вместе превышают определенный порог, способность организма сопротивляться оказывается ослабленной и индивид заболевает или умирает. Термины «порог» и «подверженность» часто используются при обсуждении мультифакториального наследования. Порог подразумевает наличие резкого качественного различия: за этим порогом на шкале подверженности располагаются пораженные индивиды. Хотя понятие порога полезно для моделей мультифакториального наследования, вряд ли он на самом деле физически существует. Концепция подверженности подразумевает градуированный континуум возрастающей восприимчивости к заболеванию. Эта концепция сложнее аналитически, но с биологической точки зрения она, вероятно, применима к большинству ситуаций. При редких заболеваниях с простым типом наследования мутация в единичном гене нарушает его функцию. В других случаях мутация приводит к трудностям лишь при особых обстоятельствах, как, например, при моногенно детерминированных реакциях на лекарства. Большинство признаков, однако, настолько сложны, что прямой анализ всех факторов оказывается практически невозможным, поскольку в подверженность вовлечено, вероятно, множество разных генов. Мы опять оказываемся в ситуации «черного ящика» - генетический анализ проще провести статистическими, нежели биологическими методами. Генетические предсказания на таком сложном уровне должны основываться на нескольких предположениях: 1) подверженность к заболеванию распределена более или менее нормально, и распределение имеет одну моду; 2) подверженность обусловлена большим числом генов, действующих аддитивно, и каждый из них вносит равный вклад; 3) когда подверженность превышает определенный порог, индивид заболевает или у него появляются нарушения. Хотя порог может быть четко определен, в большинстве случаев существует некоторая пороговая область, внутри которой от дополнительных средовых факторов будет зависеть, заболеет индивид или нет. Если опи- 250 3. Формальная генетика человека сывать это в тех терминах, которые были введены выше, то можно сказать, что наследуемость меньше единицы (рис. 3.58). Очевидно, что эта модель слишком упрощает реальность, но она может быть полезна для понимания природы ряда широко распространенных заболеваний и пороков развития. Эксперименты на животных. В экспериментальной генетике млекопитающих наследование некоторых признаков, например полидактилии у морской свинки (Райт [961]), объясняли пороговым эффектом. Скрещивались две линии: представители одной имели три пальца на задних конечностях (норма), у животных другой линии было четыре пальца (морфологический вариант). В поколении F1 обнаружено лишь несколько особей с четырьмя пальцами, тогда как во втором поколении (в потомстве скрещивания F1 х F1 этот признак имелся примерно у четверти всех особей. Генетический анализ предположительно указывал на то, что скрещиваемые линии различаются по набору диаллельных систем с аддитивным эффектом четырех локусов: любое животное могло нести максимум восемь и минимум ноль положительных аллелей. При скрещивании двух гомозиготных линий (8 х 0) (рис. 3.59) гетерозиготное поколение F 1должно иметь четыре положительных аллеля. Такой генотип приводит к четырехпалым задним конечностям только в исключительных случаях. В поколении F2 (F1 x F1) присутствуют все комбинации положительных аллелей, что дает непрерывное распределение. В этом случае было принципиально показано, что аддитивное действие генов на самом деле может быть связано с
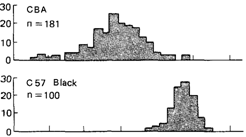
пороговым проявлением (рис. 3.59). В другом примере удалось продемонстрировать не только дискретное, но и непрерывное фенотипическое выражение количественно варьирующей подверженности. Грюнеберг (1952) [690, 691] анализировал такую систему у мыши. В инбредной линии СВА у многих особей отсутствует третий молярный зуб: у 133 из 744, т.е. по крайней мере у одной мыши из пяти. Однако в черной линии С57 этот моляр почти всегда имеется. Скрещивание двух линий (СВА х С57) обнаружило, что тип наследования не является простым, несмотря на то что признак (зуб присутствует или отсутствует) обнаруживает четко выраженное
3. Формальная генетика человека 251 альтернативное проявление. Даже у животных линии СВА с лишним зубом его размер был в среднем намного меньше, чем в черной линии С57 (рис 3.60). Следовательно, у животных линии СВА размер зуба варьирует непрерывно вплоть до определенного минимального порогового размера. Ниже этого порога зуб не формируется вовсе. Грюнеберг назвал это явление «квазинепрерывной изменчивостью». Сам по себе порог не очень четкий, правильнее говорить о некоторой пороговой области. Мультифакториальность генетической системы очевидна лишь при сопоставлении явных различий между двумя линиями и в скрещиваниях между ними. Внутри генетически однородной линии СВА изменчивость обусловлена только средовыми факторами. Предпринимались неоднократные попытки продемонстрировать непрерывно распределенную подверженность и дискретные пороги у людей (см., например [619]), но в большинстве случаев имело место лишь дискретное проявление: «поражен» или «не поражен». Чтобы установить характер внутрисемейного распределения признака с пороговым проявлением в общем случае, рассмотрим теоретическую модель. Дата добавления: 2015-12-16 | Просмотры: 775 | Нарушение авторских прав |