|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Стадии гаметогенеза1. Стадия размножения. Клетки, из которых в последующем образуются мужские и женские гаметы, называются сперматогониями и овогониями соответственно. Они несут диплоидный набор хромосом 2n2c. На этой стадии первичные половые клетки многократно делятся митозом, в результате чего их количество существенно возрастает. Сперматогонии размножаются в течение всего репродуктивного периода в мужском организме. Размножение овогоний происходит главным образом в эмбриональном периоде. У человека в яичниках женского организма процесс размножения овогоний наиболее интенсивно протекает между 2 и 5 месяцами внутриутробного развития. К концу 7 месяца большая часть овоцитов переходит в профазу I мейоза. Если в одинарном гаплоидном наборе количество хромосом обозначить как n, а количество ДНК — как c, то генетическая формула клеток в стадии размножения соответствует 2n2c до синтетического периода митоза (когда происходит репликация ДНК) и 2n4c после него. 2. Стадия роста. Kлетки увеличиваются в размерах и превращаются в сперматоциты и овоциты I порядка (последние достигают особенно больших размеров в связи с накоплением питательных веществ в виде желтка и белковых гранул). Эта стадия соответствует интерфазе I мейоза. Важное событие этого периода — репликация молекул ДНК при неизменном количестве хромосом. Они приобретают двунитчатую структуру: генетическая формула клеток в этот период выглядит как 2n4c. 3. Стадия созревания. Происходят два последовательных деления — редукционное (мейоз I) и эквационное (мейоз II), которые вместе составляют мейоз. После первого деления (мейоза I) образуются сперматоциты и овоциты II порядка (с генетической формулой n2c), после второго деления (мейоза II) — сперматиды и зрелые яйцеклетки (с формулой nc) с тремя редукционными тельцами, которые погибают и в процессе размножения не участвуют. Так сохраняется максимальное количество желтка в яйцеклетках. Таким образом, в результате стадии созревания один сперматоцит I порядка (с формулой 2n4c) дает четыре сперматиды (с формулой nc), а один овоцит I порядка (с формулой 2n4c) образует одну зрелую яйцеклетку (с формулой nc) и три редукционных тельца. Отмеченные выше различия в ходе овогенеза и сперматогенеза имеют определенный биологический смысл, связанный с разным функциональным назначением мужских и женских гамет (помимо переноса генетической информации). Накопление в цитоплазме яйцеклетки большого количества запасных питательных веществ необходимо, так как на этой «базе» осуществляется развитие дочернего организма из оплодотворенного яйца. Неравномерное клеточное деление при овогенезе и обеспечивает формирование крупной яйцеклетки. Функция же сперматозоидов заключается в отыскании яйцеклетки, проникновении в нее и доставке своего хромосомного набора. Их существование кратковременно, а поэтому нет необходимости в запасании большого количества веществ в цитоплазме. А поскольку сперматозоиды в массе гибнут в процессе поиска яйцеклетки, их образуется огромное количество. Центральное событие в процессе гаметогенеза — редукция диплоидного набора хромосом (в ходе мейоза) и формирование гаплоидных гамет. 4. Стадия формирования, или спермиогенеза (только при сперматогенезе). В результате этого процесса каждая незрелая сперматида превращается в зрелый сперматозоид (с формулой nc), приобретая все структуры, ему свойственные. Ядро сперматиды уплотняется, происходит сверхспирализация хромосом, которые становятся функционально инертными. Комплекс Гольджи перемещается к одному из полюсов ядра, формируя акросому. К другому полюсу ядра устремляются центриоли, причем одна из них принимает участие в формировании жгутика. Вокруг жгутика спирально закручивается одна митохондрия. Почти вся цитоплазма сперматиды отторгается, поэтому головка сперматозоида ее почти не содержит. 5. 6. Сперматогене́з — развитие мужских половых клеток (сперматозоидов), происходящее под регулирующим воздействием гормонов. Одна из форм гаметогенеза. Сперматозоиды развиваются из клеток-предшественников, которые проходят редукционные деления (делениямейоза) и формируют специализированные структуры (акросома, жгутик и пр.). В разных группах животных сперматогенез различается. У позвоночных животных сперматогенез проходит по следующей схеме: вэмбриогенезе первичные половые клетки — гоноциты мигрируют в зачаток гонады, где формируют популяцию клеток, называемых сперматогониями. С началом полового созревания сперматогонии начинают активно размножаться, часть из них дифференцируется в другой клеточный тип — сперматоциты I порядка, которые вступают в мейоз и после первого деления мейоза дают популяцию клеток, называемых сперматоцитами II порядка, проходящих впоследствии второе деление мейоза и образующих сперматиды; путём ряда преобразований последние приобретают форму и структуры сперматозоида в ходе спермиогенеза.
Сперматогенез у человека[править | править вики-текст]
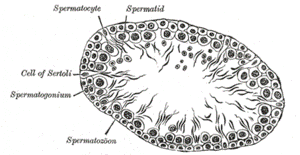
Схема поперечного сечения семенного канальца Сперматогенез у человека в норме начинается в пубертатном периоде (около 12 лет) и продолжается до глубокой старости. Продолжительность полного сперматогенеза у мужчин составляет примерно 73—75 дней[1]. Один цикл зародышевого эпителия составляет приблизительно 16 дней[2] Сперматозоиды образуются в яичках, а именно в извитых семенных канальцах. Стенка семенного канальца делится базальной мембраной на люминальную и адлюминальную стороны. На люминальной стороне расположены клетки Сертоли (сустентоциты) и предшественники половых клеток (сперматогонии, сперматоциты I и II порядков и сперматиды). Сперматогонии, лежащие непосредственно на базальной мембране извитых семенных канальцев, проходят несколько последовательных стадий митотического деления. Общее количество сперматогоний в яичке мужчины составляет около 1 млрд. Различают две основные категории сперматогоний: А и В. Сперматогонии А, которые делятся митотически, сохраняют способность к делению и поддерживают свою популяцию. Остальные дифференцируются в сперматогоний В, которые «эвакуируются» клеточными контактами сустентоцитов (образуют под основанием половой клетки новый контакт и резорбируют старый). Сперматогония В делится митотически, дифференцируясь в сперматоцит I порядка, вступающий в мейоз. В результате первого деления мейоза образуются две дочерние клетки сперматоциты второго порядка, каждый из которых содержит гаплоидный набор (23 у человека)d-хромосом [3]. Вторичные сперматоциты расположены ближе к просвету канальца. Во втором делении мейоза образуются две сперматиды. Таким образом, в результате деления одной сперматогонии образуются четыре сперматиды, каждая из которых обладает гаплоидным набором хромосом. В ходе сложного процесса спермиогенеза сперматиды дифференцируются в зрелые сперматозоиды. Дифференцирующиеся сперматиды лежат в углублениях плазматической мембраны клеток Сертоли. При спермиогенезе комплекс Гольджи формирует акросому, содержащую протеолитические ферменты, которые при контакте с яйцеклеткой растворяют участок её блестящей оболочки (zona pellucida). Сложный процесс сперматогенеза регулируется гонадотропными гормонами гипофиза и стероидными гормонами яичка. После полового созревания гипоталамусначинает выделять гонадотропный рилизинг-гормон, под влиянием которого гипофиз секретирует фолликулостимулирующий гормон (ФСГ), стимулирующий развитие и функционирование клетки Сертоли и лютеинизирующий гормон (ЛГ), стимулирующий клетки Лейдига к выработке тестостерона. Тестостерон оказывает воздействие на развитие клеток Сертоли, а также на предшественники половых клеток (в ассоциации с андроген-связывающим белком, выделяемым клетками Сертоли). Секреторная активность гипофиза регулируется клетками Сертоли и клетками Лейтига. Тестостерон, выделяемый клетками Лейдига подавляет активность гипофиза к выработке ЛГ и ФСГ. Ингибин и эстрадиол, образующийся в клетках Сертоли, подавляют гипофиз к выработке ФСГ и клетки Лейдига к выработке тестостерона. Морфофункциональное состояние яичка регулируется гормонами аденогипофиза — ФСГ и ЛГ, причём уровень гормонов постоянный, имеются лишь незначительные колебания. Вопрос№9 Характеристика овогенеза. Овогенез - процесс развития женских половых клеток - включает три периода: размножение, рост и созревание. Вопрос№10 1.Аутосомно-доминантный тип наследования: 2.Аутосомно-рецессивный тип наследования: 3.Наследование сцепленное с Х хромосомой, если ген, контролирующий проявления признака, - рецессивный: а. Мужчины наследуют чаще, чем женщины 4.Наследование сцепленное с Х хромосомой, если ген, контролирующий проявления признака, - доминантный: а. Мужчины наследуют реже, чем женщины 5.Наследование сцепленное с У хромосомой:
Типы и варианты наследования признаков . Наследование — это внешнее выражение наследственности в том смысле, что механизмы наследственности обусловливают правила (закономерности) наследования. Это хорошо иллюстрируют результаты исследований Г. Менделя, который в своих опытах наблюдал внешние проявления генетической связи между растениями разных поколений и сформулировал три правила независимого наследования признаков. От правил он пришел к формулировке законов наследственности, в которых отразил закономерности организации и динамики генетического материала — дискретность и аллельное состояние генов, генетическая «чистота» гамет и т. д. Выделяют несколько основных типов наследования признаков, каждый из которых имеет варианты. Учитывая особенности генетического контроля качественных и количественных признаков, в отношении отдельных признаков различают моногенное и полигенное наследование, о которых шла речь ранее. Моногенное наследование бывает аутосомным или тепленным с полом. Аутосомное и сцепленное с Х-хромосомой наследование может быть доминантным и рецессивным. Названные типы и варианты соответствуют правилам наследования отдельно взятых менделирующих признаков. Закономерности соотносительного наследования нескольких признаков описываются такими типами, как независимое, частично сцепленное и полностью сцепленное наследование. Тины и варианты наследования, свойственные другим организмам, обнаружены и у человека и могут быть проиллюстрированы родословными людей. Аутосомно-доминантный вариант наследования обусловливается передачей в ряду поколений доминантного аллеля гена, локализующегося в аутосоме, для которого характерны определенные черты, проявляющиеся в родословных: 1) при достаточном числе потомков признак обнаруживается в каждом поколении; 2) редкий признак наследуется примерно половиной детей; 3) потомки мужского и женского пола наследуют признак одинаково часто; 4) оба родителя в равной мере передают признак детям. Отмеченные черты характеризуют, например, наследование признака «шерстистых волос» появившихся несколько поколений тому назад в одной т норвежских семей. Аутосомно-рецессивный вариант наследования обусловливается передачей в ряду поколений рецессивного аллеля гена, локализующегося в аутосоме. Типичные черты этого варианта, выявляемые путем анализа родословных, сводятся к следующему: 1) даже при достаточном числе потомков признак может отсутствовать в поколении детей, но появляется в поколении внуков; 2) признак может развиться у детей при отсутствии его у обоих родителей; среди детей, рождаемых в таких семьях, признак обнаруживается в 25% случаев; 3) признак наследуется всеми детьми, если оба родителя имеют его; 4) признак развивается у 50% детей, если он обнаруживается у одного супруга; 5) признак наследуется потомками мужского и женского пола одинаково часто. Перечисленные особенности проявляются в исследовании альбинизма (рис. 42). Особенностью аутосомно-доминанта о го наследования при неполном доминировании и кодоминировании служит менее выраженное развитие признака у потомков — гетерозигот по сравнению с доминантными гомозиготами. Так, изменения скелета у лиц с доминантным аллелем брахидактилии (короткопалость) более выражены у гомозигот, чем у гетерозигот. У людей, гетерозиготных по локусу синтеза р-полипептида гемоглобина HbА/HbS), содержание нормального гемоглобина составляет 55—75%. Особенности вариантов наследования признаков, сцепленных с полом, отражают факт локализации соответствующих генов в половых хромосомах X и Y. Напомним, что эти хромосомы за редким исключением не имеют гомологичных локусов, поэтому по большинству генов Х-хромосомы клетки мужского организма гемизиготны. Вариант наследования при локализации гена в Y -хромосоме (голандрическое наследование) отличается прямой передачей признака от отца всем сыновьям и соответствующим обнаружением этого признака в каждом поколении у потомков мужского пола. В настоящее время неизвестны признаки, для которых голандрическое наследование было бы доказано безоговорочно. Можно думать, что так наследуется признак «волосатых ушей» (гипертрихоз края ушной раковины), который проявляется в том, что в юношеском возрасте на ушной раковине вырастает пучок длинных жестких волос.«Описана родословная, согласно которой мужчины пяти поколений имели этот признак. Особенности варианта наследования, сцепленного с полом, при локализации гена в Х-хромосоме зависят от гемизиготности соответствующих локусов у мужчин и функционально-генетической Неполное доминирование, меняющаяся экспрессивность и неполная пенетрантность аллеля, зависимость развития признака от гормонального фона, возраста организма и действие факторов окружающей среды способны затушевать картину. Дополнительные трудности в определение варианта наследования вносят генокопии — случаи, когда одно и то же состояние признака развивается под контролем разных генов. Так, фенилкетонурия возникает при дефиците синтеза как фенилаланингидроксилазы. Независимое наследование отличается случайным комбинированием признаков родителей у потомков. Оно свойственно признакам, которые контролируются генами из негомологичных хромосом, а также находящимися в одной хромосоме на расстоянии более 50 морганид. У человека независимо наследуются, например, способность ощущать вкус фенилтиомочевины и выделение в слюну эритроцитарных антигенов системы групп крови АВО. При полностью сцепленном наследовании соответствующие признаки родителя развиваются у потомка всегда одним блоком. Так наследуются признаки, контролируемые генами из одной хромосомы, которые расположены настолько близко, что это исключает кроссинговер. Полностью сцепленное наследование наблюдается для таких признаков, как синтез полипептидов р- и 6-гемоглобина человека. При частичном сцепленном наследовании соответствующие признаки родителей воспроизводятся у части потомков совместно, а у части — независимо. Такое наследование наблюдается для генов одной группы сцепления, расстояние между которыми в хромосоме допускает регулярный кроссинговер. Так, у человека гены, контролирующие выделение в слюну антигенов системы групп крови АВО и системы Lutheran, расположены в одной хромосоме на расстоянии 15 морганид. Эти признаки наследуются по частично сцепленному типу. Вопрс№11
Дата добавления: 2015-12-16 | Просмотры: 1509 | Нарушение авторских прав |