|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Краткая характеристика темы. При анализе наследования целого ряда признаков причиной отклонения от менделевских закономерностей в распределении генотипов и фенотипов в потомстве является
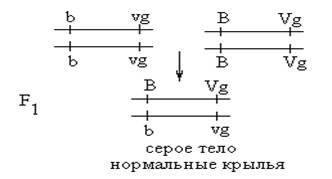
При анализе наследования целого ряда признаков причиной отклонения от менделевских закономерностей в распределении генотипов и фенотипов в потомстве является сцепление генов. Явление сцепленного наследования было открыто Морганом в 1912 году в экспериментах с плодовой мушкой Drosophilla melanogaster. Для скрещивания использовались мухи, различающиеся по двум парам признаков: серое (В)- чёрное (в) тело; нормальные (Vg) - рудиментарные (vg) крылья. Серое тело и нормальные крылья- доминантные признаки. Опыты Моргана можно представить следующей схемой: Р: ♀ черное тело х ♂ серое тело рудиментарные крылья нормальные крылья
Все мухи первого поколения оказались единообразными, согласно I закону Менделя, Далее были проведены 2 серии анализирующих скрещиваний.
В первой серии анализаторами были самки: Р: ♀ черное тело х ♂ серое тело рудиментарные крылья нормальные крылья
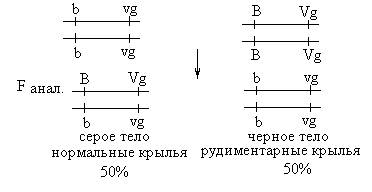
В потомстве анализирующего скрещивания появилось только 2 фенотипических класса, полностью повторяющие родительские сочетания признаков. На основании этой серии опытов Морган сделал вывод, что гены В и Vg находятся в одной хромосоме. Гены, находящиеся в одной хромосоме, наследуются вместе, или сцепленно, и называются группой сцепления. Число групп сцепления в организме равно гаплоидному набору хромосом. Во второй серии анализирующего скрещивания в качестве анализатора использовались самцы: Р: ♀ серое тело х ♂ черное тело нормальные крылья рудиментарные крылья
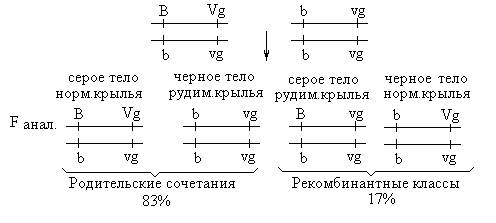
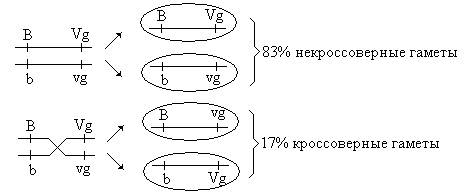
Таким образом, в потомстве второй серии анализирующих скрещиваний появились 4 фенотипических класса, но с разной вероятностью. Два фенотипических класса (BVg и bvg) повторяли родительские сочетания признаков и составляли 83%, а 17% особей имели новые сочетания признаков (Bvg и bVg). Появление рекомбинантных классов в потомстве Морган объяснил кроссинговером. Кроссинговер – обмен гомологичными участками гомологичных хромосом – происходит в профазе I мейотического деления. Результатом кроссинговера является образование кроссоверных гамет с новым сочетанием генов и появление рекомбинантных классов в потомстве. Если между генами, относящимися к одной группе сцепления (хромосоме), происходит кроссинговер, говорят о неполном сцеплении этих генов. Процент кроссоверных потомков равен проценту кроссоверных гамет, образуемых особью. Таким образом, самки F1 дали 4 типа гамет:
Частота кроссинговера не превышает 50%, поэтому кроссоверных потомков всегда меньше, чем некроссоверных. Полное сцепление генов, наблюдавшееся Морганом в первой серии анализирующего скрещивания, объясняется тем, что у самцов дрозофилы кроссинговера не происходит. Таким образом, существует два вида сцепления: полное и неполное. При полном сцеплении гены, локализованные в одной хромосоме, всегда наследуются вместе. Неполное сцепление генов характеризуется нарушением групп сцепления в результате процесса кроссинговера. При этом, чем дальше гены расположены в хромосоме друг от друга, тем чаще между ними происходит кроссинговер. Исходя из этого положения, ученик Моргана Стертевант предложил использовать частоту кроссинговера в качестве единицы измерения расстояния между генами:
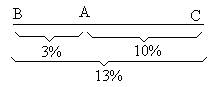
Расстояние между генами выражается в сантиморганидах (сМ). 1 сантиморганида = 1% кроссинговера. Расстояние 5 сМ означает, что особь образует 5% кроссоверных гамет и 95% некроссоверных гамет. На основании работ Моргана и его школы были сформулированы основные положения хромосомной теории наследственности: 1. Материальной основой наследственности являются хромосомы. 2. Гены расположены в хромосоме в линейном порядке. 3. Гены, находящиеся в одной хромосоме, наследуются вместе и называются группой сцепления. Это положение известно как закон Моргана. 4. Число групп сцепления равно гаплоидному набору хромосом. 5. Сила сцепления генов в хромосоме обратно пропорциональна расстоянию между ними. Принцип определения расстояния между генами на основе расчета частоты кроссинговера был положен в основу составления генетических карт хромосом у разных видов организмов. Генетическая карта представляет собой схему линейного расположения генов в хромосоме с указанием расстояния между ними в сМ от одного из концов хромосомы, принятого за начало отсчета (0,0 сМ). Генетическую карту строят постепенно, исходя из данных о частоте кроссинговера между двумя последовательно расположенными наиболее близкими между собой генами. В качестве пояснения рассмотрим следующий пример: гены А, В и С сцеплены. В результате серии анализирующих скрещиваний установлено, что АВ = 3%, АС = 10%, ВС = 13%. Расположить ген В относительно генов А и С.
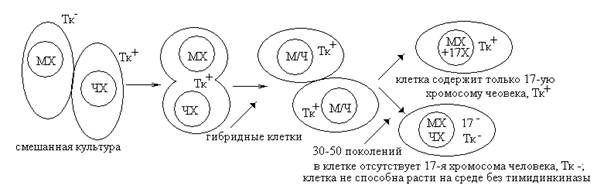
Ген В локализуется слева от гена А. На основании таких экспериментов в настоящее время для многих организмов составлены генетические карты хромосом. К сожалению, в силу целого ряда причин (невозможность произвольного скрещивания, малое число потомков и т.д.) этот метод неприменим для составления генетических карт человека. При анализе групп сцепления человека наиболее часто используют метод соматической гибридизации клеток и современные молекулярно-генетические методы картирования (метод ДНК-зондов). Большинство генов человека, контролирующих определенные этапы метаболического пути (ферментативные гены), картировано с помощью метода соматической гибридизации. Для этой цели используют чаще всего эмбриональные фибробласты человека и опухолевые клетки мыши. Гибриды соматических клеток получают, стимулируя их слияние воздействием инактивированного вируса Сендай или полиэтиленгликоля. Такие соматические гибриды содержат хромосомы мыши (МХ) и хромосомы человека (ЧХ). Соматические гибридные клетки, как правило, нестабильны и при следующих делениях имеют тенденцию к утрате многих хромосом человека. Поэтому через определенное число делений можно получить клетки, утратившие или сохранившие какую-либо одну конкретную хромосому человека. Используя подходящую селективную систему, можно отобрать клетки с определенной ферментативной активностью и локализовать ген этого фермента в конкретной хромосоме. Таким образом, например, было определено место локализации гена тимидинкиназы (Тк) в 17 хромосоме человека. Схему опыта можно представить следующим образом: (в качестве селективной системы использовать клетки мыши, лишенные тимидинкиназной активности).
Вывод: ген, отвечающий за синтез тимидинкиназы, локализован в 17-ой хромосоме человека.
Дата добавления: 2015-12-16 | Просмотры: 706 | Нарушение авторских прав |