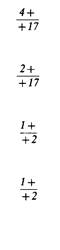В первом приближении это предположение нашло подтверждение в рассмотренных опытах по изучению генной конверсии. Напомним, что половина всех наблюдаемых генно-конверсионных событий в локусе arg 4 сопровождалась рекомбинацией фланкирующих маркеров. Это можно рассматривать, как генетическое подтверждение процесса изомеризации структур Холлидея (рис. 14.1, Ж, 3) и дополнительное указание на то, что структуры этого типа выступают в качестве интермедиатов при генетической рекомбинации у эукариот. В действительности «горизонтальное» расщепление структуры Холлидея, не сопровождающееся рекомбинацией фланкирующих маркеров, но приводящее к закреплению гетеродуплексных участков в дочерних молекулах ДНК, проявляется подобно двойному кроссинговеру и характеризуется высокой отрицательной интерференцией даже в отсутствие репарации неправильных пар нуклеотидов (см. первую диаграмму на рис. 14.9, Б).
Дополнение 14.1. Гетероаллельная конверсия
в локусе arg 4 дрожжей
Данные, приведенные в таблице 14.1, отражают процессы гетероаллельной конверсии, наблюдаемые в асках, образуемых четырьмя различными диплоидными генотипами Saccharomyces cerevisiae, каждый
из которых гетерозиготен по каким-либо двум из четырех гетероаллелей, отмеченных на генетической карте (рис. 148) Кроме того, каждый из диплоидов гетерозиготен по фланкирующим маркерам,
Таблица 14.1. Связь генной конверсии в локусе arg 4 с рекомбинацией фланкирующих маркеров
Диплоид
Генотип arg 4
Число асков
Конвертируемые
аллели
Число конверсии
3 1 плюс 1 3
Фланкирующие маркеры
Число конверсии, сопровождающихся рекомбинацией фланкирующих маркеров
BZ34
4
pet 1—17
17 4-17
42 5
4—thr1 pet1 — thr1
BZI40
2
pet1—17
17 2-17
7 28
2— thr1 pet1 — thr1
3 14
Х841
1
pet1 — 2
2 1-2
14 19
1— thr1 pet1 — thr1
8 8
Х901
1
c—2
2 1-2
1—thr1 c— thr1
3 4
Всего
По: Hurst D, Fogel S, Mortimer R (1972) Proc Nat Acad Sei USA, 69, 101