в частности pet1 и thr1, за рекомбинацией которых при мейозе можно вести наблюдение. Например, диплоид BZ34 характеризуется наличием маркеров arg 4-4 и thr1 на одной и pet1 и arg 4-1 7 на другой гомологичной хромосоме:
Для 690 асков, образуемых этим диплоидом, наблюдалось восемь случаев конверсии arg 4-4, семь из которых сопровождались рекомбинацией маркеров pet1 и arg 4-1 7, фланкирующих arg 4-4. В приведенных данных не делается различий между конверсией от arg 4-4 к дикому типу и конверсией от дикого типа к arg 4-4, обнаруживаемых в асках с распределением аллелей 1:3 и 3:1. Кроме того, были зарегистрированы 42 случая конверсии arg 4-1 7, из которых 19 сопровождались рекомбинацией фланкирующих маркеров arg 4-4 и thr1. И наконец, в 5 случаях наблюдалась двойная конверсия arg 4-4 и arg 4-17, при этом в 3 случаях имела место рекомбинация фланкирующих маркеров pet1 и thr1. В таблице 14.1 аналогичным· образом представлены также результаты, полученные при изучении конверсии для трех других диплоидов.
На основании данных, приведенных в таблице 14.1, можно сделать три важных заключения. Во-первых, около половины (73/140) всех конверсионных событий связаны с рекомбинацией фланкирующих маркеров. Во-вторых, конверсия представляет собой замену одного исходного аллеля на другой, участвующий в скрещивании. Так, аллель arg 4-4 дикого типа переходит при конверсии в мутантный аллель arg 4-4, а мутантный arg 4-4 -в соответствующий аллель дикого типа. То есть конверсия не является случайным событием в том смысле, что не приводит к конверсии одного гетероаллеля в другой произвольный аллель. И в-третьих, соконверсия двух гетероаллелей при скрещивании происходит чаще, чем этого можно было бы ожидать от двух независимых событий. Так, частота конверсии для arg 4-4 составляет 0,012 (8/690), а для arg 4-1 7-0,061 (42/690). Если бы два этих события были совершенно независимы, то частота совместной конверсии составляла бы 0,00073 (0,012 χ 0,061). В то же время наблюдаемая частота совместной конверсии этих маркеров в действительности достигает 0,0072 (5/690). Этот и аналогичные ему расчеты, сделанные на основании данных таблицы 14.1 для других пар гетероаллелей arg4, приведены в таблице 14.2.
Таблица 14.2. Ожидаемая и наблюдаемая частоты совместной конверсии в дрожжевом гене arg 4
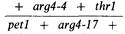 Для 690 асков, образуемых этим диплоидом, наблюдалось восемь случаев конверсии arg 4-4, семь из которых сопровождались рекомбинацией маркеров pet1 и arg 4-1 7, фланкирующих arg 4-4. В приведенных данных не делается различий между конверсией от arg 4-4 к дикому типу и конверсией от дикого типа к arg 4-4, обнаруживаемых в асках с распределением аллелей 1:3 и 3:1. Кроме того, были зарегистрированы 42 случая конверсии arg 4-1 7, из которых 19 сопровождались рекомбинацией фланкирующих маркеров arg 4-4 и thr1. И наконец, в 5 случаях наблюдалась двойная конверсия arg 4-4 и arg 4-17, при этом в 3 случаях имела место рекомбинация фланкирующих маркеров pet1 и thr1. В таблице 14.1 аналогичным· образом представлены также результаты, полученные при изучении конверсии для трех других диплоидов.
Для 690 асков, образуемых этим диплоидом, наблюдалось восемь случаев конверсии arg 4-4, семь из которых сопровождались рекомбинацией маркеров pet1 и arg 4-1 7, фланкирующих arg 4-4. В приведенных данных не делается различий между конверсией от arg 4-4 к дикому типу и конверсией от дикого типа к arg 4-4, обнаруживаемых в асках с распределением аллелей 1:3 и 3:1. Кроме того, были зарегистрированы 42 случая конверсии arg 4-1 7, из которых 19 сопровождались рекомбинацией фланкирующих маркеров arg 4-4 и thr1. И наконец, в 5 случаях наблюдалась двойная конверсия arg 4-4 и arg 4-17, при этом в 3 случаях имела место рекомбинация фланкирующих маркеров pet1 и thr1. В таблице 14.1 аналогичным· образом представлены также результаты, полученные при изучении конверсии для трех других диплоидов.