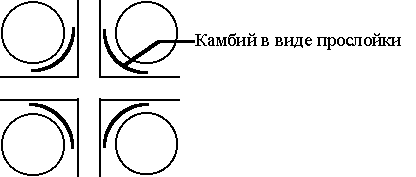|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Первичное строение корняЕсли разрезать корень несколько выше в зоне проведения, то уже на малом увеличении микроскопа хорошо выделяется две зоны. Широкая первичная кора, занимающая большую часть поперечного сечения среза и сравнительно узкий центральный осевой цилиндр. Такое соотношение тканевых комплексов? характерная особенность корня, в стебле все по- другому: большую часть поперечного сечения стебля занимает стела. Наружный слой первичной коры называется экзодермой. Он приходит на смену отмирающему и разрушающемуся волосконосному слою, остатки которого можно обнаружить на некоторых препаратах. Экзодерма - это первичная покровная ткань корня. Клетки экзодермы плотно сомкнуты и обычно опробковевают. У большинства растений экзодерма однослойна, хотя у отдельных растений, например, у ириса-касатика, она многослойна. Под экзодермой залегает мощный слой клеток паренхимы первичной коры. Обычно это тонкостенные клетки многогранной или округлой формы, расположенные без особого порядка. Между паренхимными клетками всегда имеются межклетники. По межклетникам циркулируют газы, необходимые для дыхания клеток корня. У некоторых водных и болотных растений система межклетников развита настолько сильно, что первичная кора превращается в аэренхиму. Таким образом, мощность воздухоносной системы напрямую зависит от аэрации субстрата. В паренхимных клетках коры протекает энергичный метаболизм. Поэтому нередко в них можно обнаружить друзы кристаллов, как отходы метаболизма, или запасные вещества, например, крахмальные зерна. Внутренний слой коры представлен эндодермой. Это своеобразная внутренняя пограничная ткань, отделяющая первичную кору корня от центрального осевого цилиндра. Эндодерма сплошным кольцом плотно сомкнутых клеток окружает стелу. У однодольных растений клетки эндодермы имеют весьма своеобразные подкововидно-утолщенные оболочки, пропитанные суберином. Жироподобное вещество - суберин непроницаемо для воды и газов, поэтому эндодерма прекрасно выполняет свою пограничную функцию. Однако изоляция стелы от первичной коры не может быть сплошной, поэтому среди клеток эндодермы обнаруживаются отдельные клетки, не имеющие характерных утолщений оболочки. Они работают как своеобразные ворота, по которым вещества могут перемещаться из коры в стелу и обратно и называются пропускными клетками. Теперь самая сложная часть корня? центральный осевой цилиндр или стела. Сложная, потому что развитие проводящих тканей необходимо рассмотреть в динамике. Из гистогена - плеромы образуется две первичные меристемы - перицикл и прокамбий. Обычно перицикл тонким слоем окружает прокамбий.
Из прокамбия образуются все остальные ткани стелы корня. Наиболее важно рассмотреть особенности формирования проводящих тканей: флоэмы и ксилемы. Первой в корне закладывается флоэма. Закладывается она снаружи прокамбиального тяжа, как говорят, экзархно. Это означает, что на периферии тяжа группы клеток прокамбия превращаются в элементы флоэмы. Далее во флоэму превращаются все более внутренне расположенные слои прокамбия, т.е. ее развитие идет центростремительно.
Ксилема по времени закладывается позднее и тоже снаружи прокамбиального тяжа, т.е. экзархно. Как и флоэма она развивается по направлению к центру прокамбиального тяжа. Однако в своем развитии ксилема обгоняет флоэму и первой достигает центра корня. В результате, в стеле корня формируется весьма своеобразная конструкция: многолучевая звезда ксилемы, между лучами которой располагаются массивы флоэмы. Звезда ксилемы может иметь разное количество лучей: два, и тогда ее называют диархной; четыре - тетрархная, пять - пентархная и т.д. Таким образом, обе проводящие ткани в корне закладываются и развиваются одинаково: экзархно и центростремительно. В стебле развитие проводящих тканей происходит по-другому. Однако между однодольными и двудольными растениями на заключительной стадии формирования корня первичного строения наблюдаются существенные различия. Так, у однодольных растений весь прокамбий полностью (нацело) расходуется на образование проводящих тканей - флоэмы и ксилемы. Поэтому флоэма и ксилема плотно прилегают друг к другу, между ними не остается никакой меристематической прослойки, "никакого зазора". У двудольных же растений между лучами ксилемы и массивами флоэмы остается неизрасходованная прослойка прокамбия. Со временем у клеток этой прослойки повторно пробуждается меристематическая активность и прослойка начинает работать как вторичная меристема, т.е. превращается в камбий. Первичную структуру приобретают корни всех растений. У большинства однодольных растений и у папоротникообразных она сохраняется на протяжении всей жизни. Именно поэтому однодольные растения являются наиболее удобными объектами для изучения корня первичного строения.
У двудольных же растений в корне происходят вторичные изменения. Вторичная структура развивается за счет функционирования вторичных меристем: камбия и феллогена (пробкового камбия).
Дата добавления: 2015-02-02 | Просмотры: 1172 | Нарушение авторских прав |
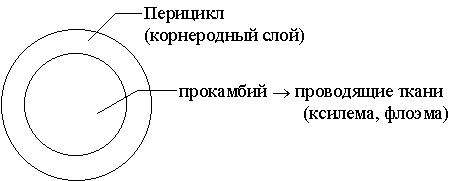 Перицикл дает начало боковым корням, которые закладываются эндогенно и, пронзая мощный слой первичной коры, постепенно выходят наружу. На этом основании перицикл называют корнеродным слоем.
Перицикл дает начало боковым корням, которые закладываются эндогенно и, пронзая мощный слой первичной коры, постепенно выходят наружу. На этом основании перицикл называют корнеродным слоем.