|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Поглощение минеральных веществ корневой системой растений.Корневая система растений поглощает из почвы как воду, так и питательные вещества. Эти процессы взаимосвязаны, но осуществляются на основе разных механизмов. Многочисленные исследования показали, что почти никогда раствор питательных солей не поступает в растение в неизменной концентрации. Из очень разбавленных растворов соли поглощаются быстрее, чем вода, и раствор становится еще более разбавленным. Наоборот, из концентрированных растворов растение берет больше воды, чем солей, и раствор становится еще более концентрированным. Растения поглощают вещества избирательно, в результате соотношение поглощенных веществ обычно оказывается иным, чем в питательном растворе (табл. 19). В тканях актинидии китайской концентрация фосфора в несколько тысяч раз, а калия и азота в десятки раз превышает содержание этого элемента в почвенном растворе. Эти данные свидетельствуют также о том, что поглощение веществ идет не только избирательно, но и против градиента концентрации. Процесс поглощения веществ делят на два этапа: поступление ионов в свободное пространство корня; транспорт их в протопласты клеток. При помещении корней растений на раствор катионного красителя, например метиленового синего, можно наблюдать, что в первые же 3 мин из раствора поглощается примерно половина того количества, которое поглотится за длительное время. Дальнейшее поглощение будет идти очень медленно (часами). Действие на корневую систему дыхательных ядов или пониженной температуры полностью устраняет последующее медленное поглощение вещества из раствора. При переносе корней в воду или раствор соли наблюдается обратная картина: быстрое выделение красителя в первые несколько минут и последующий медленный его выход из ткани. Причем количественно выход красителя в раствор превышает выход в воду. Первоначальное быстрое поглощение веществ осуществляется в клеточных стенках и является адсорбцией, а быстрое выделение — десорбцией. Больший выход красителя в раствор соли свидетельствует об обменном характере адсорбции. Та часть объема корня, где происходят процессы обменной адсорбции, получила название кажущегося свободного пространства (КСП). Термин «кажущееся» означает, что объем этого пространства зависит от объекта и природы растворенного вещества. КСП включает межмолекулярные пространства в толще клеточных стенок и на поверхности плазмалем- мы и клеточных стенок и занимает 5—10 % объема корня. Высокая адсорбирующая способность обусловлена отрицательно заряженными карбоксильными группами пектиновых веществ (полигалактуроновых кислот), мозаично заряженными белками и доннановским электропотенциалом на границе водной среды и катионообменника. Эти факторы уже на первом этапе обеспечивают некоторую избирательность поглощения веществ, несущих заряд, так как клеточные стенки более активно связывают катионы, чем анионы. Из-за высокой плотности отрицательных фиксированных зарядов в клеточной стенке происходит первичное концентрирование катионов в пространстве, непосредственно прилегающем к плазмалемме. В почве корень контактирует с водной фазой (почвенным раствором) и с частицами почвы, которые также являются преимущественно катионообменниками (почвенный поглощающий комплекс). Катионы и анионы поступают в клеточные стенки ризодермы как непосредственно из почвенного раствора, так и благодаря контактному обмену с частицами почвенного поглощающего комплекса. Оба эти процесса связаны с обменом катионов Н+ на катионы окружающей среды и HC Способность к обменной адсорбции определяется обменной емкостью корня. Она зависит от химического состава корневых выделений и клеточных стенок и поддерживается транспортом веществ через цитоплазматическую мембрану и дальнейшим их перемещением внутрь корня. Ионы и различные соединения могут преодолевать мембрану по электрохимическому градиенту за счет простой диффузии путем растворения в липидной фракции мембраны или через гидрофильные поры (ионные каналы), а также за счет облегченной диффузии гидрофильных веществ с помощью липофильных переносчиков. Возможно также поступление ионов против электрохимического градиента с затратой метаболической энергии при участии активных переносчиков (насосов). Особую роль в плазмалемме растительных клеток играет Н+ насос (протонная помпа), создающий электрический и химический (∆рН) градиенты ионов Н+. Электрический потенциал ионов Н+ (мембранный потенциал) может быть использован на транспорт катионов по электрическому градиенту против концентрационного. В свою очередь, ∆рН служит энергетической основой для переноса через мембрану анионов в симпорте с ионами Н+, возвращающимися через мембрану по концентрационному градиенту. Аналогично Н+ могут функционировать и HC Именно мембранное строение клеток обеспечивает избирательность поглощения веществ растением. Поддержание структуры мембран, их обновление и функционирование активных механизмов транспорта ионов являются энергозависимыми процессами. Поэтому существует тесная связь поглотительной деятельности корня с дыханием. При действии дыхательных ядов и низких температур происходит только первая фаза поглощения веществ — их проникновение в кажущееся свободное пространство, дальнейшее поступление веществ в протоплазму отсутствует из-за ограниченности энергетических ресурсов клетки. Наиболее активно поглощают вещества клетки зоны растяжения и корневых волосков. Корневые волоски не только позволяют корню осваивать значительно больший объем почвы, но и обладают более высокой поглотительной активностью. В опытах О. О. Лялина и И. С. Ахмедова установлено, что молодые, только начинающие рост корневые волоски имеют на порядок большую проницаемость для ионов, чем расположенные рядом с ними без- волосковые клетки. По мере старения корневого волоска и затухания его роста число ионных каналов на единицу клеточной поверхности снижается быстрее, чем возрастает сама поверхность. Корневой волосок интенсивно поглощает ионы, пока он растет. Наличие большого количества точек роста в корневой системе обеспечивает ее высокую поглотительную активность. Дата добавления: 2015-12-15 | Просмотры: 1880 | Нарушение авторских прав |
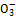 или анионов органических кислот на анионы минеральных веществ
или анионов органических кислот на анионы минеральных веществ