|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
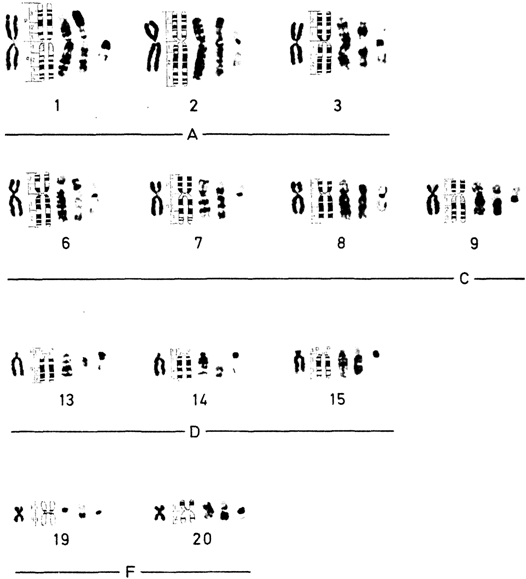
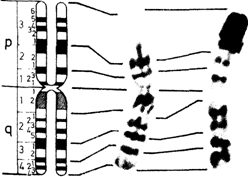
Нормальный кариотип человека в метафазе митозаСтандартное окрашивание. Хромосомы располагаются и нумеруются в зависимости от их длины. Согласно Денверской классификации, предложено нумеровать пары хромосом от 1 до 23 (1960). Патау (1960) [471] показал, однако, что в некоторых группах хромосомы нельзя однозначно классифицировать, и предложил идентифицировать пары на основании относительной длины и положения центромеры. В соответствии с этой последней характеристикой различают метацентрические, субметацентрические и акроцентрические хромосомы. Патау предложил разбить 23 пары на восемь групп от А до G. Это предложение было принято в качестве альтернативной процедуры. Все три пары метацентрических хромосом группы А можно идентифицировать. В группе Е обычно легко выделить хромосому 16, а хромосомы 17 и 18 поддаются идентификации, только если препараты высокого качества. Y-хромосому обычно можно отличить от других хромосом группы G. Все остальные хромосомы групп В, С (включая Х-хромосому), D, F и G не идентифицируются. Важным параметром является центромерный индекс, который отражает отношение (в %) длины короткого плеча к длине всей хромосомы. Дифференциальное окрашивание. Кариотип человека представлен на рис. 2.8-2.10, использованы различные методы. В настоящее время каждую хромосому можно идентифицировать. На рис. 2.11 изображены схема распределения G- и Q-сегментов и их нумерация. Отдельные хромосомы, а также их наиболее часто встречающиеся «нормальные» варианты описываются следующим образом [392]. Индивидуальные характеристики хромосом человека. Группа А (1-3). Большие, метацентрические и субметацентрические хромосомы. Хромосома 1 – самая большая метацентрическая хромосома. Центромера расположена посередине, центромерный индекс 48-49. В проксимальной части длинного плеча вблизи центромеры часто обнаруживается «вторичная перетяжка», что в ряде случаев приводит к удлинению длинного плеча (рис. 2.12). Растянутый сегмент по сравнению с остальной частью хромосомы может выглядеть очень тонким, а по сравнению со сверхспирализованными метафазными – «недоспирализованным» (uncoiler). Признак «недоспирализации», как и другие индивидуальные особенности хромосомной морфологии, обнаруживается во всех соматических и в половине половых клеток, т. е. наследуется как простой доминантный признак. Локус uncoiler- 1, определяющий этот признак, был использован для картирования локуса Даффи на хромосоме 1 (разд. 3.4). При окрашивании Q-методом вторичная перетяжка флуоресцирует слабо, при использовании G-метода она выглядит как плотный сегмент. Самой большой субметацентрической хромосомой является хромосома 2 с центромерным индексом 38-40. Радиоавтографическое исследование с 3Н-тимидином показало, что хромосома 2, особенно проксимальные районы обоих плеч, реплицируется относительно поздно. Хромосома 3 с центромерным индексом 45-46 почти на 20% короче хромосомы 1 и, следовательно, легко идентифицируется. При окрашивании Q-методом в проксимальном районе ее длинного плеча часто выявляется ярко флуоресцирующий сегмент. Интенсивность флуоресценции значительно варьирует у разных индивидов, но постоянна во всех клетках для одного и того же хромосомного варианта. Группа В (4 и 5). Большие субмета- 46 2. Хромосомы человека
центрические хромосомы (центромерный индекс 24-30) не различаются между собой без радиоавтографии или дифференциального окрашивания. Согласно данным радиоавтографических исследований, хромосома 4 является поздно реплицирующейся по всей своей длине, в то время как в хромосоме 5 поздно реплицируется только короткое плечо. Рисунки распределения Rи G-сегментов у этих хромосом совершенно различны. Группа С (6-12). Хромосомы среднего 2. Хромосомы человека 47
размера, субметацентрические. При стандартном окрашивании Х-хромосому нельзя отличить от других хромосом этой группы. Хромосомы 6, 7, 8, 11 и 12 являются относительно субметацентрическими, их центромерный индекс 27-35. В хромосоме 9 часто обнаруживают вторичную перетяжку в проксимальной части длинного плеча. Все эти хромосомы легко идентифицируются с помощью Q- и G-окрашивания. Вторичная перетяжка хромосомы 9 не окрашивается ни акрихином, ни красителем Гимза. Хро- 48 2. Хромосомы человека
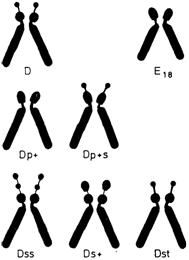
мосомы 11 и 12 обнаруживают очень сходный рисунок сегментации, что наводит на мысль об их общем происхождении и эволюции (они содержат локусы лактатдегидрогеназы А и В соответственно, общее происхождение которых предполагается на основании биохимических данных). Однако хромосома 11 заметно более метацентрическая, чем хромосома 12. В противоположность другим хромосомам этой группы Х-хромосома значительно варьирует по длине. В целом она сходна с самыми длинными из С хромосом. Центромерный индекс высокий, но довольно вариабельный. В клетках женщин одна из двух Х-хромосом реплицируется в поздней S-фазе, когда репликация других С-хромосом (за исключением ряда коротких сегментов) уже завершена. Важно, что поздним является не только завершение, но также и начало репликации ДНК. Группа D (13-15). Эти хромосомы акроцентрические по форме, сильно отличаются от всех других хромосом человека. Центромерный индекс около 15-наименьший в кариотипе человека. Все три пары содержат спутник. Короткое плечо этих хромосом обнаруживает сильную межхромосомную вариабельность. Длина проксимальных участков коротких плеч варьирует, спутники могут отсутствовать, а могут быть очень большими, могут ярко флуоресцировать, а могут и не давать флуоресценции. В некоторых случаях наблюдаются двойные (тандемные) спутники. Длинные плечи трех хромосом четко различаются по Q- и G-сегментам. Для выявления вариантов в группе D—G сравнивают длины короткого плеча этих хромосом с длиной короткого плеча хромосомы 18 в той же клетке. Плечо считают длинным (ph +), если оно такой же длины, как короткое плечо хромосомы 18, и очень длинным, если оно длиннее короткого плеча этой хромосомы. Большие 2. Хромосомы человека 49
спутники обозначают (ps +), двойные спутники (pss), укороченное короткое плечо со спутниками или без них (ph —). Частота гетероморфизма гомологов в этой группе составляет 3,7% (8 из 216) в препаратах после дифференциального окрашивания и 2,3% (411 из 24400) в стандартных препаратах [422] (рис. 2.14). Группа Е (16-18). Относительно короткие метацентрические или субметацентрические хромосомы. Хромосома 16 имеет центромерный индекс около 40. В среднем ее длина составляет чуть более одной трети
50 2. Хромосомы человека
длины хромосомы 1, но обнаруживает значительную изменчивость. В длинном плече примерно в 10% случаев выявляется вторичная перетяжка. Длина проксимального G-сегмента варьирует в зависимости от выраженности этой перетяжки. Хромосома 18 примерно на 5-10% короче хромосомы 17 и имеет более короткое длинное плечо (у хромосомы 17 центромерный индекс составляет 31 по сравнению с 26 у хромосомы 18). Хромосома 17 реплицируется рано, хромосома 18-поздно. Группа F (19-20). Эти две хромосомы имеют центромерный индекс в пределах 36-46. В стандартных препаратах они выглядят одинаково, но при дифференциальном окрашивании резко различаются. Группа G (21 и 22). У этих маленьких акроцентрических хромосом центромерный индекс варьирует в пределах 13-33. Они легко различаются по рисунку сегментации. Изменчивость их коротких плеч так же значительна, как и в хромосомах группы D. Здесь классифицируют такие же варианты, как и в группе D (рис. 2.14). Флуоресценция спутников и коротких плеч может быть слабой, умеренной и сильной, так же как и интенсивность окрашивания при использовании G-метода. В выборке из 2444 новорожденных 3,5% обнаруживают удлиненные короткие плечи. Другие варианты, такие, как гигантские спутники, удлиненные или укороченные короткие плечи, встречаются намного реже. По данным некоторых исследователей, общая частота вариантов хромосом группы G составляет 1,8% по препаратам с дифференциальным окрашиванием и 1,6% в стандартных препаратах. Короткие плечи хромосом группы D и G содержат ядрышковый организатор и специфично окрашиваются методом серебрения. Y-хромосома обычно (но не всегда) больше, чем хромосомы группы G, и хроматиды ее длинного плеча, как правило, лежат параллельно одна другой. Этим она отличается от хромосом группы G, у которых хроматиды длинных плеч часто образуют широкий угол. Центромера видна менее четко, спутники отсутствуют, размер длинного плеча сильно варьирует, и некоторые варианты его длины наследуются. 2. Хромосомы человека 51 Центромерный индекс варьирует от 0 до 26 (в среднем ~ 16). При окрашивании акрихином обнаруживается довольно изменчивый ярко флуоресцирующий дистальный участок длинного плеча. Во многих случаях находят два сильно флуоресцирующих сегмента, реже – три. В популяционных исследованиях частота выраженных вариантов размеров длинного плеча Y-хромосомы составляет 5,6% (в выборке из 2444 новорожденных). В большинстве случаев Yхромосома была удлиненной; у 5% новорожденных она оказалась длиннее Fхромосомы, у 0,33% - длиннее хромосомы 18; однако в 0,25% образцов была обнаружена очень маленькая Y-хромосома. Хроматин [201, 516]. В интерфазных ядрах дистальный интенсивно флуоресцирующий участок длинного плеча Y-хромосомы выявляется как яркое пятно диаметром 0,3-1,0 мкм. На рис. 2.15 показан Y-хроматин в эпителиальных клетках, гранулоцитах, больших лимфоцитах и в сперматозоидах. Измерения хромосом. Измерения митотических хромосом сопряжены с определенными трудностями, так как положение центромеры не всегда можно определить достаточно точно. Парижская конференция (1971) [468] разработала рекомендации относительно измерений хромосом. В табл. 2.1 представлены усредненные данные о размерах митотических хромосом человека. Гетероморфизм хромосом. Морфология отдельных хромосом не всегда одинакова у разных индивидов. Гетероморфизм особенно выражен в отношении размера спутничной области акроцентрических хромосом, длины Y-хромосомы, особенно ее гетерохроматического участка, и «вторичных перетяжек» хромосом 1 и 9. Характерен он и для гетерохроматических сегментов других хромосом (о гетерохроматине см. разд. 2.3.14). При анеуплоидии гетероморфизм гомологов по гетерохроматическим районам можно использовать для выяснения происхождения данной хромосомы от одного из родителей (разд. 5.1.2.3).
Во многих хромосомах обнаруживаются фрагильные (ломкие) участки, т. е. участки, подверженные хромосомным и хроматидным разрывам (разд. 2.2.2). Такие разрывы относительно просто индуцировать удалением фолиевой кислоты из питательной среды [390а]. Недавно обнаружена ассоциация фрагильного участка в дистальном районе длинного плеча Х-хромосомы с характерной формой умственной отсталости (разд. 8.2.1.2). Высокоразрешающее дифференциальное окрашивание. Хромосомы в профазе и прометафазе конденсированы не столь сильно, как метафазные хромосомы. При обработке культуры лимфоцитов метотрексатом (для частичной синхронизации клеточного цикла) можно накопить достаточное число клеток, находящихся в профазе и прометафазе. Сокращение времени инку- 52 2. Хромосомы человека
2. Хромосомы человека 53 Таблица 2.1. Относительная длина (в процентах от длины гаплоидного набора аутосом) и центромерный индекс (отношение длины короткого плеча к общей длине хромосомы х 100). Хромосомы окрашены орсеином или методом Гимза 9 и предварительно идентифицированы с помощью Q-метода. (Парижская конференция, 1971 [468].)
бации с колцемидом позволяет избежать сильной конденсации. В препаратах таких хромосом отдельные сегменты, выявляемые стандартными методами, можно подразделить на субсегменты. Степень разрешения зависит от стадии, на которой клетки были зафиксированы. Некоторые авторы описывают свыше 2000 сегментов [550]. Обычно в поздней профазе можно увидеть 800-1200 сегментов (рис. 2.16). Хотя этот метод не заменяет стандартный, используемый при рутинной диагностике, однако он полезен для более точной идентификации точек разрывов и мелких аберраций, например в случае наследуемых сбалансированных и несбалансированных транслокаций или особенно в цитогенетике опухолей.
Электронно-микроскопическая картина хромосом [490, 517]. Чтобы выявить тонкую структуру хромосом человека, были использованы многочисленные методы электронной микроскопии. Современные модели организации генетического материала эукариот будут обсуждаться в разд. 2.3, здесь же достаточно сказать, что данные электронной микроскопии не противоречат модели, предполагающей, что хроматин состоит из сверхспирализованных нитей, причем имеется несколько порядков спирализации (рис. 2.17). Обнаружено три типа хроматиновых фибрилл: фибриллы первого типа имеют диаметр 250 Å, фибриллы второго типа – 100 Å и третьего - только 30-50 Å. Имеются довольно убедительные доказательства того, что фибриллы этого последнего типа представляют собой генетически активный хроматин. Двойная спираль чистой ДНК имеет диаметр ~ 20 Å, следовательно, фибриллы 30-50 Å соответствуют диаметру нити ДНК вместе с белками (гистонами и негистонами). Фибриллы диаметром 100 Å отражают, по-видимому, вторичную спирализацию фибрилл 30-50 Å, а нити 250 Å могут отражать третичный уровень спирализации. В метафазной хромосоме эти «третичные спирали» могут иметь примерно такую укладку, как указано на рис. 2.17. Примерно девять фибрилл 250 Å, вероятно, каким-то образом связаны вместе, и два таких пучка образуют различимую 54 2. Хромосомы человека
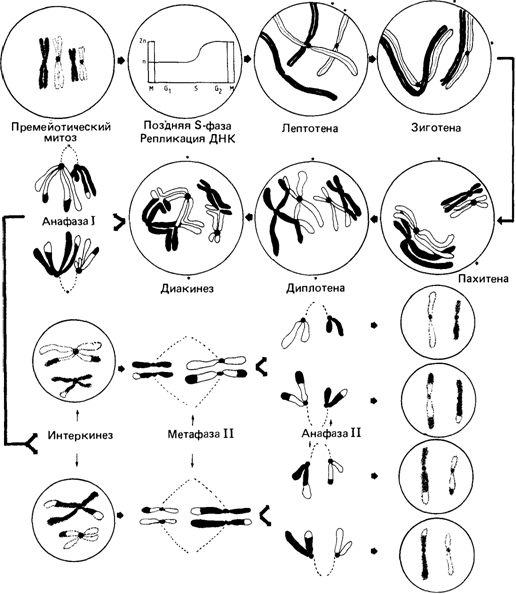
на электронно-микроскопических изображениях спиральную структуру, характерную для каждой хромосомы [490]. В отдельных препаратах обнаруживаются остатки мембраны, предположительно ядерной. Некоторые исследователи считают этот факт доказательством того, что интерфазные хромосомы в разных точках прикреплены к мембране. Следует учесть, однако, что процесс приготовления препаратов для электронной микроскопии хромосом включает целый ряд процедур, и потому трудно решить, существуют ли эти или другие структуры in vivo или они являются попросту артефактами. Мейоз Биологическая функция мейоза. Благодаря митозу поддерживается постоянство числа хромосом в ряду клеточных поколений. В отличие от митоза мейотический процесс обеспечивает уменьшение (редукцию) диплоидного числа хромосом (46 у человека) наполовину до гаплоидного (23 у человека). При оплодотворении в результате слияния двух гаплоидных половых клеток в зиготе восстанавливается диплоидное число 46, которое сохраняется во всех последующих митотических делениях. В мейозе расхождение гомологичных хромосом в разные половые клетки происходит случайно, что увеличивает генетическую изменчивость. Соматические клетки являются диплоидными (2n), они содержат обе гомологичные хромосомы одной пары, в то время как половые клетки гаплоидны (n) и несут только один гомолог из каждой пары. Последний цикл регулярного синтеза ДНК происходит в интерфазе непосредственно перед первым мейотическим делением и предшествует фазам мейоза, показанным на рис. 2.18. Первое деление мейоза. Профаза I. На этой стадии становятся видимыми длинные хромосомные нити (лептотена), затем происходит конъюгация (спаривание) гомологичных хромосом, которая часто начинается с теломерных районов (зиготена). Точный молекулярный механизм конъюгации хромосом еще не известен. Две конъюгированные гомологичные хромосомы (называемые на этой стадии «бивалентом») формируют на субмикроскопическом уровне характерную двойную структуру, так называемый синаптонемальный комплекс (рис. 2.19). К моменту завершения конъюгации хромосомы вследствие спирализации становятся короче и толще (пахитена). В каждом биваленте обнаруживается продольная щель и становятся видимыми расположенные бок о бок четыре хроматиды (диплотена). В то время как сестринские хроматиды остаются спаренными, несестринские - разделяются. На этой стадии несестринские хроматиды соединяются между собой в некоторых точках, образуя фигуру, напоминающую греческую букву χ. Такие фигуры получили название «хиазм». Метафаза I. Хромосомы располагаются в экваториальной плоскости, а их центромерные районы оттянуты к полюсам. Гомологичные хромосомы начинают разделяться, но еще удерживаются в участках хиазмообразования, особенно часто – в дистальных районах. Анафаза I. Начинается «терминализация» хиазм, т. е. они перемещаются к концам хромосом и затем исчезают. Гомологичные хромосомы окончательно разделяются и перемещаются к противоположным полюсам. Образуются дочерние ядра (интеркинез). Второе деление мейоза. В принципе, это митотическое деление удвоенного гаплоидного набора хромосом. Как указывалось выше, мейоз начинается после завершения последнего цикла репликации ДНК, в результате чего количество генетического материала в ходе первого деления остается учетверенным (2x2 гомологичных хромосом), но после завершения второго деления оно распределяется по четырем половым клеткам. Второй важный аспект мейоза состоит в случайном распределении негомологичных хромосом, благодаря чему существует большое число возможных комбинаций хромосом в разных половых клетках. При наличии у человека 23 пар хромосом число возможных комбинаций в одной гамете составляет 223 = 8 388 608. Число возможных комбинаций хромосом в потомстве данной пары родителей состав- 2. Хромосомы человека 55
56 2. Хромосомы человека
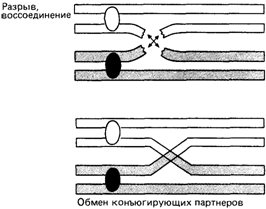
ляет 223 х 223, а в действительности еще больше – за счет кроссинговера (перекреста), происходящего во время конъюгации гомологичных хромосом. Морфологическим проявлением кроссинговера являются хиазмы. Каждая хиазма соответствует одному событию кроссинговера, в котором участвуют две несестринские хроматиды (рис. 2.20). Одно время активно дебатировался вопрос о том, происходит ли кроссинговер во время последнего цикла репликации ДНК посредством механизма «выбора копии» или уже после синтеза ДНК путем разрыва и последующего крестообразного воссоединения несестринских хроматид в гомологичных сайтах (рис. 2.21). Эта альтернатива теперь, по-видимому, разрешена в пользу гипотезы «обменов». Например, в профазе I наблюдается так называемый внеплановый синтез ДНК, который вполне может отражать процесс воссоединения концов при кроссинговере. Молекулярные механизмы рекомбинации не являются специфической проблемой генетики человека. Они обстоятельно обсуждаются в руководствах по молекулярной генетике. Сперматогенез. С наступлением половой зрелости сперматоциты мужчины постоянно претерпевают мейотические деления. После второго мейотического деления происходит плотная упаковка ДНК и митохондрий и завершается формирование спермиев, которые приобретают способность активно двигаться. Препараты хромосом на стадии сперматогониальных митозов или на стадии мейотического деления можно получить из материала биопсии тестикул, удаленных при хирургической операции. Хромосомы на стадии диакинеза в 2. Хромосомы человека 57
мейозе у мужчины показаны на рис. 2.22. Гомологи еще тесно прилежат один к другому в их теломерных районах, в то время как центромерные районы уже начали перемещаться к полюсам. Половой бивалент четко отличается от всех остальных благодаря тому, что Х- и Y-хромосомы ассоциируют «конец в конец» и хиазмы в нем не обнаруживаются. Во время пахитены половой бивалент начинает конденсироваться раньше других и находится в «половом пузырьке». Часть района короткого плеча Х-хромосомы и короткое плечо Y-хромосомы конъюгируют (рис. 2.23). Гиб- 58 2. Хромосомы человека
ридизационные эксперименты с ДНК-зондами показали, что эти районы структурно гомологичны [502]. В случае свободной рекомбинации генов, локализованных в гомологичных сегментах Х- и Y-хромосом, их поведение не должно было бы отличаться от поведения аутосомных генов. Такие «псевдоаутосомные» Х- и Y-сцепленные гены действительно были идентифицированы [315а; 488а]. Холдейн (1936) [372], учитывая возможность редкого кроссинговера между Х- и Y-хромосомами, предположил существование частичного сцепления с полом тех генов человека, которые локализованы в гомологичном для Х- и Y-хромосом сегменте. Однако удовлетворительные доказательства такого частичного сцепления с полом у человека пока не получены. Более того, локусы стероид-сульфатазы и эритроцитарного антигена Xg, расположенные очень близко к псевдоаутосомному району Х-хромосомы, сегрегируют в соответствии с классическим Х-сцепленным наследованием. Среднее число хиазм на клетку и размах изменчивости по этому показателю приведены в табл. 2.2. Некоторые биваленты могут содержать несколько хиазм, свыше пяти и даже шесть. Исходя из числа хиазм, генетическая длина (разд. 3.4) генома человека составляет около 25,5 морганиды у мужчин; у женщин эта величина больше, но точные оценки еще не получены [450]. Домовая мышь – единственное млекопитающее, кроме человека, для которого получены достоверные оценки, – имеет геном длиной 16,2-19,2 морганиды [88]. Оогенез. У всех млекопитающих оогенез сильно отличается от сперматогенеза. Общая схема представлена на рис. 5.13 (разд. 5.1.3.3). На рис. 2.24 и 2.25 приведена схема цитологических процессов. Ооциты полностью формируются уже на поздней эмбриональной стадии. После диплотены клетка переходит в стадию диктиотены, для которой характерна морфология хромосом типа «ламповых щеток». В этой стадии мейоз останавливается на долгие годы. После рождения большинство ооцитов дегенерирует. В процессе полового созревания некоторые ооциты начинают расти, заканчивают первое мейотическое деление и вступают в профазу II и затем в метафазу II. В это же время начинается овуляция. Мейоз завершается только после оплодотворения. Вокруг женских и мужских гаплоидных наборов хромосом образуется ядерная мембрана, и зигота теперь содержит два «пронуклеуса». Эта стадия особо чувствительна к нарушениям, выз-
2. Хромосомы человека 59
ванным, например, мутагенными агентами (разд. 5.2.1). Несколько часов спустя два пронуклеуса сливаются, образуя диплоидное ядро, и зигота начинает делиться путем обычных митозов. Исследование мейотических хромосом в оогенезе сопряжено с большими трудностями. Было опубликовано лишь несколько удовлетворительных микрофотографий (рис. 2.26). Анализ генетического сцепления показывает, что кроссинговер у женщин происходит чаще, чем у мужчин (разд. 3.4), следовательно, и хиазм у женщин должно быть больше. У женщин только одна из четырех клеток - продуктов мейоза развивается в ооцит, три другие формируют полярные тельца, которые в норме не оплодотворяются. 60 2. Хромосомы человека
2. Хромосомы человека 61
Обычно считают, что вероятность для хромосомы оказаться в полярном тельце не зависит от ее генетических особенностей. Данные о сохранении стандартных сегрегационных вероятностей для большинства генных мутаций (50:50, 25:75 и т. д.) свидетельствуют, что это допущение справедливо. Однако имеются и исключения (разд. 3.1.4): в случае структурных аберраций хромосом возможно неслучайное расхождение нормальных и аберрантных гомологов в полярные тельца. Половые различия в мейозе. Две основные особенности отличают мейоз у мужчин и женщин: 1. У мужчин все четыре клетки, образующиеся в результате мейотического деления, развиваются в зрелые гаметы, в то время как у женщин только одна из них становится зрелым ооцитом, остальные дегенерируют. 2. У мужчин мейоз следует непосредственно за серией митотических делений; он завершается, когда сперматиды на- 62 2. Хромосомы человека
чинают трансформироваться в зрелые спермин. У женщин мейоз начинается на очень ранних стадиях эмбрионального развития, и ему предшествует намного меньше оогониальных митотических делений. После этого мейотический процесс прерывается на длительный период и завершается только после оплодотворения. Эти различия важны для генетики человека. То обстоятельство, что только одна из четырех клеток развивается в зрелый ооцит, а три полярных тельца почти (или совсем) не имеют цитоплазмы, дает возможность этому ооциту передать новой зиготе полный набор цитоплазматических компонентов, таких, как митохондрии и информационные РНК (разд. 4.7.1). Эти различия в клеточной кинетике, вероятно, обусловливают разницу между мужчинами и женщинами в частоте трисомий, с одной стороны, и точковых мутаций – с другой (разд. 5.1 и 5.2). Дата добавления: 2015-12-16 | Просмотры: 1003 | Нарушение авторских прав |