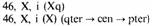|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
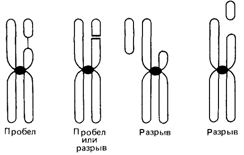
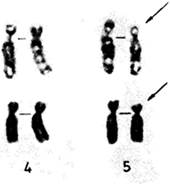
Кариотипы u клинические синдромыПервые наблюдения синдрома Дауна. Как только трисомия 21 была идентифицирована как причина синдрома Дауна, естественно возник вопрос о том, у всех ли больных имеется эта трисомия. Если не у всех, то исключения могли бы представлять большой интерес. Так как риск мейотического нерасхождения, как уже тогда было известно, увеличивается с возрастом матери и поскольку единичное нерасхождение должно вести к появлению только одного пораженного потомка, исключения следовало искать среди пораженных детей молодых матерей, а также в семьях с двумя или более больными. Полани и сотр. (1960) [479] исследовали трех таких больных с синдромом Дауна. У одной девочки, первого ребенка 21-летней матери и 23-летнего отца, они обнаружили 46 хромосом. Было найдено четыре хромосомы группы G. Однако одна хромосома из группы D имела удлиненное короткое плечо. Авторы предположили, что дополнительная хромосома 21 была транслоцирована на короткое плечо D-хромосомы. Очень скоро это предположение подтвердилось при исследовании семейных случаев. Две здоровые матери трех больных с синдромом Дауна и их общая бабка имели только 45 хромосом и только 3 стандартные хромосомы группы G. Однако одна из хромосом группы D (исследователи предположили, что это хромосома 15) имела удлиненное короткое плечо. Если это плечо содержит материал отсутствующей хромосомы 21, тогда кариотип этих женщин является сбалансированным, т.е. весь генетический материал диплоидного набора присутствует. С другой стороны, у некоторых из их потомков имеется хромосома с транслокацией, включающей большую часть материала хромосомы 21. Фактически у таких детей имеется трисомия 21 и возникает синдром Дауна, несмотря на то что формально число хромосом у них стандартное. Такой кариотип является несбалансированным. Примерно в то же время была описана первая транслокация G/G [354]. Вскоре после этого при исследовании первого мейотического деления у гетерозиготного носителя сбалансированной транслокации был обнаружен тривалент, т.е. фигура, состоящая из трех хромосом, и это послужило четким доказательством того, что нестандартная хромосома, обнаруженная в этих семьях, действительно несет транслокацию [373]. Частота транслокационного синдрома Дауна. Транслокация при синдроме Дауна объясняет много семейных случаев, но не все. Стандартная трисомия 21 может повторно возникать в одной и той же семье, указывая на наличие у родителей каких-то конституциональных факторов, предрасполагающих к нерасхождению, или мозаицизму (разд. 5.1.2). В табл. 2.3 приведены данные о частоте транслокационных случаев (наследуемых и спорадических) среди больных с синдромом Дауна для двух групп матерей: молодых и пожилых. Большинство случаев характеризуется описанными выше транслокациями D/G и G/G. Существует, однако, небольшое число реципрокных транслокаций, в которые вовлекаются другие – неакроцентрические хромосомы. Детальное обсуждение различных структурных аберраций целесообразно предварить замечаниями относительно механизмов их образования. Пробелы и разрывы. Необходимым условием возникновения структурной хромосомной перестройки любого типа является наличие в хромосоме разрыва. Если исходить из того, что ДНК представляет собой единую длинную нить, проходящую через всю хромосому, хромосомный разрыв предполагает и разрыв сахаро-фосфатного остова ДНК. В световом микроскопе бывает трудно отличить хромосомный разрыв от ахроматической (неокрашенной) области, называемой пробелом. 72 2. Хромосомы человека
Эти пробелы могут отражать как истинные разрывы, так и участки локальной деспирализации. Хромосомные разрывы часто учитывают при оценке мутационного процесса, поэтому необходимо прийти к соглашению относительно того, какие аберрации учитывать как разрывы, а какие – как пробелы. Схема, положенная в основу одного из таких соглашений, представлена на рис. 2.36. Указанные в ней отличительные признаки достаточно строгие и, вероятно, занижают количество разрывов. Разрывы и пробелы могут возникать во время интерфазы как до, так и после репликации ДНК. Если разрыв происходит до репликации, повреждение будет видно в последующей метафазе в обеих хроматидах (изохроматидный разрыв). Если событие произойдет после фазы репликации, поврежденной окажется только одна хроматида (хроматидный разрыв). Различные типы разрывов и пробелов представлены на рис. 2.37. Судьба поврежденных хромосом. Разрыв, происходящий в любом районе хромосомы и не затрагивающий центромеры, приводит к появлению укороченной хромосомы с центромерой и ацентрического фрагмента. Такой фрагмент иногда может формировать маленькое кольцо, но, будучи лишенным центромеры, чаще всего теряется в последующем митозе. Таким образом, разрыв хромосомы часто приводит к появлению клетки, лишенной хромосомного сегмента. В некоторых случаях, однако, целостность хромосомы, имеющей разрывы в двух точках, восстанавливается ферментами репарации. Механизмы такого воссоединения концов в настоящее время известны [456]. Если концы хромосомных фрагментов воссоединятся друг с другом удачно, то и хромосома,' и клетка будут снова интактными. Действительно, исследования при заболеваниях, связанных с недостаточностью репаративных ферментов, показывают, что подобные события могут происходить многократно во многих тканях. В других случаях концы хромосомных фрагментов могут воссоединиться в точках разрыва других хромосом как гомологичных, так и негомологичных (при условии, что два разрыва происходят в пределах относительно короткого отрезка времени и достаточно близко друг от друга.) 2. Хромосомы человека 73
Это приводит к образованию хромосомных перестроек различного типа. Внутрихромосомные перестройки (внутренние обмены). В пределах одной хромосомы могут произойти разрывы в двух разных участках, и фрагмент между точками разрыва, перевернувшись, может вновь соединиться с хромосомой. Такая перестройка (инверсия) не приводит к нарушениям в митозе, особенно если разрыв произошел в фазе G 1. Она может быть обнаружена методами дифференциального окрашивания. В тех случаях, когда инверсия не затрагивает центромеру, она называется парацентрической, если же точки разрыва находятся по обе стороны от центромеры, такую инверсию называют перицентрической. Гетерозиготы по инверсиям не очень редки в популяциях человека (рис. 2.38). Инверсии могут создавать затруднения в конъюгации гомологичных хромосом в мейозе и приводить к частичной элиминации некоторых типов половых клеток у гетерозигот по инверсиям (рис. 2.39). У гомозигот таких затруднений нет. Инверсии (особенно перицентрические), несомненно, играли важную роль в филогении высших приматов (разд. 7.2.1). Другой тип внутренних обменов представляют кольцевые хромосомы (рис. 2.40). Перестройка этого типа возникает при утрате обоих теломерных участков хромосомы (как ацентрических фрагментов) и 74 2. Хромосомы человека последующем воссоединении открытых концов. Судьба кольцевой хромосомы в митозе зависит от того, как завершилось воссоединение концов сестринских хроматид. Если во время репликации ДНК обмен между сестринскими нитями в точках разрыва не происходит, то кольцо, удваиваясь, образует два отдельных кольца, каждое со своей центромерой. Такие кольцевые хромосомы проходят через митоз без затруднений. Один обмен между сестринскими нитями ведет к образованию большого кольца с двумя центромерами. Дицентрическая структура обычно разрушается в наступающем митозе. Два обмена могут привести к образованию двух колец, «сцепленных» друг с другом подобно звеньям цепи. Детали различных вариантов представлены на рис. 2.40. Иногда хроматидные разрывы и образование колец происходят в фазе G2, и тогда в отдельной клетке наблюдается картина, показанная на рис. 2.41.
Межхромосомные перестройки (внешние обмены). Во многих случаях воссоединение открытых концов затрагивает разные хромосомы как гомологичные, так и негомологичные. Если разрыв происходит в фазе G 1, то воссоединение обычно завершается в той же фазе G1 (или ранней S) перед репликацией ДНК. Если каждая из перестроенных хромосом сохраняет центромеру, то такие транслокационные хромосомы могут пройти через наступающий митоз без всяких затруднений. Если одна из перестроенных хромосом приобретает две центромеры, формируется дицентрическая хромосома. В зависимости от деталей репликации она может пройти через наступающий митоз при следующих условиях: 1) если обе центромеры отойдут к одному и тому же Полюсу и 2) если репликация и сестринский хроматидный обмен между двумя центромерами не приведут к переплетению хроматид (рис. 2.42). Если разры-
2. Хромосомы человека 75
76 2. Хромосомы человека
2. Хромосомы человека 77
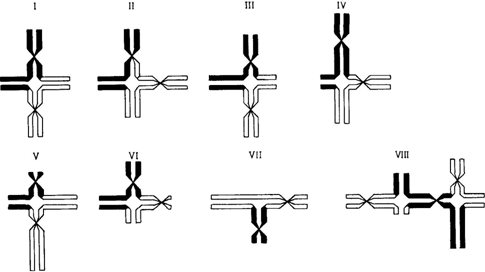
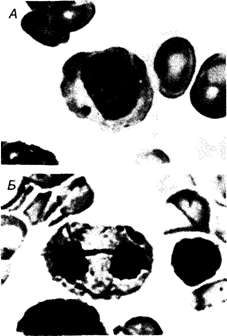
вы и воссоединения концов завершатся после репликации ДНК, то затронутой окажется только одна сестринская хроматида каждой хромосомы. Воссоединенные сестринские хроматиды еще остаются спаренными с их неповрежденными партнерами. Это ведет к межхромосомным обменам, которые обнаруживаются в первом митотическом делении после воссоединения. Различные типы этих обменов показаны на рис. 2.43. Если каждая из перестроенных хромосом сохранит центромеру (рис. 2.43; класс I, III и V), то анафазное расхождение хроматид в обеих таких хромосомах будет протекать без всяких затруднений. Однако если обе центромеры окажутся в одном и том же сегменте, то образующиеся дочерние клетки в любом случае будут анеуплоидными: либо центромеры отойдут к разным полюсам и возникнет «анафазный мост», который приведет в конце концов к разрыву, либо две центромеры отойдут к одному и тому же полюсу. В этом случае перестройка завершится только негомологичным воссоединением (рис. 2.43, классы VI, VII). Дальнейшие события откладываются до следующего митоза, в котором появляется дицентрическая хромосома. Иногда она может пройти и этот митоз. В любом случае, однако, при указанных выше условиях межхромосомные обмены, как правило, приводят к гибели клеток вследствие анеуплоидии или нарушений в митозе. В соматических тканях человека многие из этих митотических нарушений можно видеть даже в рутинных клеточных препаратах, приготовленных без использования специальных хромосомных методик. На рис. 2.44, А и Б показаны анафазные мосты и так называемые микроядра в клетках костного мозга человека. Микроядра формируются теми хромосомами (или хромосомными фрагментами), которые не связаны с митотическим аппаратом 78 2. Хромосомы человека
и не принимают участия в митозе, как остальные хромосомы. Это сопровождается преждевременной конденсацией таких хромосом и их фрагментов. В метафазных хромосомах основного ядра хроматиды имеют обычно нормальную степень конденсации, в то время как хромосомы микроядра конденсированы по типу профазных (рис. 2.45). Эти цитологические феномены полезно использовать для экспресс-оценки мутагенных агентов (разд. 5.2). Преждевременную конденсацию хромосом можно увидеть также in vitro при слиянии интерфазной клетки с другой, находящейся в предмитотической фазе [500]. Этот метод пригоден для изучения строения хромосом в интерфазном ядре. Транслокации могут привести к нарушениям и в мейозе, так как на ранних стадиях этого деления гомологичные хромосомы конъюгируют. Если в перестройке участвуют три хромосомы, как, например, у носителей сбалансированных транслокаций D/G или G/G, в метафазе I они образуют так называемые трехзвенные цепочки. На рис. 2.46 показана такая структура у носителя сбалансированной D/D-транслокации. На рис. 2. 46, А и Б представлена 2. Хромосомы человека 79 схема событий, которые происходят с этой хромосомой на стадии пахитены в процессе кроссинговера; на рис. 2.46, В – картина, которую можно ожидать в диакинезе, если каждая из двух свободных хромосом имеет один перекрест с транслокационной хромосомой. Для сравнения показана трехзвенная цепочка в том виде, как она наблюдается реально. Если в перестройку вовлечены четыре хромосомы, то образуется четырехзвенная цепочка. Такое событие ведет иногда к последующей анеуплоидии в зависимости от комбинаторики анафазных движений четырех центромер. Если две центромеры одного элемента отходят к одному полюсу и если хроматиды не переплетутся между центромерами, то деление завершится нормальной анафазой. Однако очень часто происходят дополнительные разрывы хромосом. Мейоз служит хорошим фильтром для хромосомных перестроек. Хромосомные разрывы имеют место как в соматических, так и в половых клетках. Изучение этих явлений в соматических клетках актуально с точки зрения мутационных исследований (разд. 5.2). Разрывы хромосом в половых клетках могут передаваться следующему поколению, что часто приводит к гибели зиготы на эмбриональной стадии. Однако в некоторых случаях хромосомная аберрация оказывается совместимой с постнатальной жизнью, и это приводит к рождению ребенка с хромосомным синдромом. Прежде чем перейти к анализу некоторых из этих синдромов, необходимо описать общепринятую номенклатуру кариотипа человека. Эта номенклатура была разработана группой цитогенетиков и согласована на Парижской конференции в 1971 г. [468].
Описание кариотипа человека. При описании кариотипа человека прежде всего указываются общее число хромосом и набор половых хромосом. Затем отмечается, какая хромосома лишняя, какой не хватает, а также структурно измененные. Некоторые примеры представлены в табл. 2.4.
Изменения длины вторичных перетяжек или неокрашивающихся районов следует отличать от увеличения или уменьшения длины плеча в целом, что связано с другими структурными изменениями. С этой целью специальный символ «h» помещают между символом плеча и знаками + или —
80 2. Хромосомы человека Все символы перестроек помещают перед обозначением вовлеченных хромосом, а перестроенные хромосомы (или хромосома) всегда должны быть заключены в скобки:
Номенклатура хромосомных сегментов. Каждая хромосома рассматривается как совокупность чередующихся сегментированных и несегментированных участков, границы которых указываются специальными метками. Сами сегменты и районы, которым они принадлежат, обозначаются порядковыми числами, причем центромера служит исходной точкой для цифровой схемы. При обозначении любого отдельного сегмента используются четыре метки: номер хромосомы, символ плеча, номер района и номер сегмента в пределах этого района. Например, запись 1рЗЗ означает, что речь идет о хромосоме первой пары, ее коротком плече, районе 3, сегменте 3. Сведения о номерах районов и сегментов можно почерпнуть из рис. 2.12; в табл. 2.5 приводятся рекомендуемые сокращения. Некоторые примеры иллюстрируют принцип описания.
Эти примеры должны разъяснить символы, употребляемые в нашей книге и в цитогенетических публикациях. Применение дифференциального окрашивания и высокоразрешающая сегментация требуют логического расширения номенклатуры (см. рис. 2.16). Делеционные синдромы. Индивид, гетерозиготный по делеции, является моносомиком по соответствующему району хромосомы. Де Груши и сотр. (1963) [367] первыми описали делецию del 18p-, однако делеционный синдром впервые был обна-
2. Хромосомы человека 81
ружен Леженом и сотр. в 1963 г. [418]. Ими были выявлены трое детей с делецией короткого плеча хромосомы 5 (del 5p-). Кроме обычных признаков аутосомных аномалий (общее отставание в развитии и низкий вес при рождении) у этих детей отмечалось лунообразное лицо с гипертелоризмом (широко расставленные глаза). Во внешнем облике больных не было каких-то ярких особенностей (рис. 2.47), однако их плач напоминал мяуканье кошки (cri du chat или cat cry). Существует несколько разных механизмов возникновения делеций и соответственно разные типы самих делеций: 1) истинная концевая делеция, 2) интерстициальная делеция и 3) делеция в результате транслокации. Во многих сообщениях указывается на наличие при синдроме «кошачьего крика» транслокации. На рис. 2.48 представлена часть кариотипа пробанда с хромосомой 5р-. Делетированный участок включает 5р15 и часть 5р16 сегментов. У фенотипически нормальной матери обнаружена такая же хромосома 5, но одна из хромосом 17-й пары имела лишний сегмент на длинном плече между 17q12 и 17q21. Следовательно, концевой сегмент хромосомы 5 содержится в длинном плече хромосомы 17. Случай, выявленный при помощи G-метода (рис. 2.47), иллюстрирует пример истинной концевой делеции. Внутренние обмены: парацентрические и перицентрические инверсии [401а]. Парацентрические инверсии (т.е. не вовлекающие центромеру) у человека обнаруживаются с большим трудом. Они будут обсуждаться в контексте хромосомной эволюции (разд. 7.2.1). Начиная с 60-х гг. было опубликовано много работ о предполагаемых перицентрических (т.е. захватывающих центромеру) инверсиях. У некоторых носителей таких инверсий выявлены различные аномалии типа умственной отста- 82 2. Хромосомы человека
лости или пороков развития. Фенотип других не обнаруживал каких-либо заметных отклонений, но в браках с ними регистрировались повторные спонтанные аборты. У представителей третьей группы не обнаружено вообще никаких аномалий. Следует отметить, что при использовании обычных методов окрашивания хромосом перицентрические инверсии выявляются относительно редко. С внедрением в широкую практику методов дифференциального окрашивания появились сообщения о более высокой частоте инверсий в некоторых популяциях. Довольно часто в эти перестройки вовлекается хромосома 9. Именно такая ситуация обнаружена в Финляндии [323]. При анализе кариотипов 631 жителя этой страны по различным диагностическим поводам у 9 была обнаружена перицентрическая инверсия и в 6 случаях – в хромосоме 9. Все эти б инверсий оказались идентичными. У трех пробандов инверсия была обнаружена в хромосоме 10: при этом у двух одинаковая, а у третьего отличная от них. Инверсию в хромосоме 9 может легко распознать и неспециалист (рис. 2.49), так как типичная вторичная перетяжка оказывается в этом случае не в длинном, а в коротком плече. Идентификация инверсии в хромосоме 10 требует особого опыта (рис. 2.50). При исследовании мейоза у двух пробандов с инверсией хромосомы 9 идентифицированный бивалент 9 имел нормальную морфологию, но около вторичной перетяжки не было выявлено ни одной хиазмы. Весьма вероятно, что инверсии приводят к несовершенной конъюгации и к подавлению кроссинговера, как это хорошо известно на примере других организмов, в частности у дрозофилы. Подобные инверсии можно использовать для региональной локализации генов соответствующих районов хромосомы 9 (разд. 3.4). Эти инверсии не влияют на мейотическую сегрегацию хромосом и не приводят к пренатальной гибели гетерозигот, как это следует из специальных работ (см. разд. 3.3): в браках между нормальной гомозиготой и гетерозиготой по inv (9) 25 потомков имели нормальный кариотип, 23 – были гетерозиготами. Аналогично для двух типов inv (10) суммарно это отношение оказалось равным 10:11. Еще в одном браке между двумя гетерозиготами по инверсии, оказавшимися дальними родственниками, среди детей обнаружена одна гомозигота. Такого рода наблюдения проливают свет на механизмы хромосомной эволюции (разд. 7.2.1). 2. Хромосомы человека 83 Как отмечалось выше, пробанды в этом исследовании направлялись на консультацию с диагностическими целями, поэтому вряд ли неожиданным является тот факт, что у них выявлены разнообразные аномалии. Однако эти аномалии трудно было охарактеризовать как единый синдром. Более того, среди родственников с инверсиями были и вполне нормальные в клиническом отношении. Следовательно, весьма вероятно, что инверсии в хромосомах 9 и 10 не влияют ни на фенотип носителей, ни на их плодовитость. Перицентрические инверсии обнаружены и в хромосоме 2 (рис. 2.51) [419]. В этом сообщении речь идет о трех семьях. Две из них обследованы по поводу рождения детей с пороками развития, тогда как третья - по поводу привычных выкидышей, т. е. эта выборка является сильно смещенной и возможность того, что привычные выкидыши могут быть вызваны инверсиями, исключить нельзя. Исследуя происхождение семей этих пробандов в разных странах, авторы указывают, что вряд ли данная инверсия имеет
общее происхождение. Они ссылаются на случай, где та же инверсия описана как новая мутация [383]. Более вероятным им представляется предположение о повышенной ломкости хромосомы в соответствующем сегменте. Однако ссылка на новую мутацию была привлечена до того, как стали применяться методы дифференциального окрашивания. Очень маленькие инверсии могут встречаться в отдельных популяциях довольно часто, так как скорее всего они совершенно не влияют на состояние здоровья или плодовитость. Если инверсия затрагивает протяженный участок хромосомы, то возможность нарушений в мейозе более вероятна. Однако сами носители инверсий эуплоидны, поэтому вряд ли следует ожидать у них какие-либо фенотипические аномалии. Рекомбинационная анеусомия. Встречаются семьи, в которых один из родителей, по-видимому, имеет такую же аберрацию, что и ребенок, например перицентрическую инверсию или транслокацию. При этом родитель фенотипически нормален, в то время как у ребенка обнаруживается тяжелый синдром нарушения развития. Факты такого рода можно объяснить случайным 84 2. Хромосомы человека
сочетанием в одной семье наследуемого полиморфного хромосомного варианта и каких-то нарушений развития различной этиологии. Однако в других случаях кроссинговер в участке инверсии или транслокации между аномальной хромосомой и ее нормальным гомологом может вести к появлению несбалансированных наборов хромосом в половых клетках. Такое объяснение было выдвинуто Леженом и Берже еще в 1965 г. [416], но его подтверждение получили только после появления методов дифференциального окрашивания. Впервые указанный механизм удалось продемонстрировать реально в работе [340]. Речь идет о мальчике с множественными пороками развития. На рис. 2.52 показаны хромосомы 10 этого пробанда и его матери. Можно видеть, что у матери имеется большая перицентрическая инверсия. Кроссинговер в пределах этой инверсии привел к появлению аномальной хромосомы, в результате чего ребенок оказался трисомиком по сегменту q456. Без применения метода дифференциального окрашивания все С-хромосомы (группа 6 X 12) были бы классифицированы как нормальные и кариотипы матери и ребенка рассматривались бы как идентичные. Высокоразрешающие методы позволяют выявлять такие случаи. Кольцевые хромосомы. Иная ситуация характерна для кольцевых хромосом. Поскольку образование кольца, как полагают, связано с утратой теломерных сегментов хромосомы, носители кольцевых хромосом должны напоминать носителей соответствующих делеций. Например, если в коль- 2. Хромосомы человека 85
цевую перестройку вовлечена хромосома 5р, у пробанда может наблюдаться синдром «кошачьего крика» [455]. В других случаях в зависимости от размеров делетированного участка симптомы могут быть менее выраженными. Так, например, кольцевая хромосома 13 была обнаружена у 14-месячного ребенка с умственной отсталостью и такими признаками, как микроцефалия, эпикант, широкая спинка носа, выступающие ушные раковины, микрогнатия [382]. В 85% лимфоцитов крови и в 82% фибробластов кожи выявлялось простое кольцо, идентифицированное как 13r (p11; q34). В 7% лимфоцитов и в 6% фибробластов можно было наблюдать двойное дицентрическое кольцо, которое состояло из двух хромосом 13. В 5% лимфоцитов и в 8% фибробластов кольцо отсутствовало, одна метафаза была с двумя сцепленными двойными кольцами, остальные клетки содержали другие аномалии. На рис. 2.40 показана судьба кольцевой хромосомы в митозе. В большинстве случаев кольцо реплицируется и проходит через митоз нормально. Иногда происходит один сестринский обмен и формируется двойное кольцо с двумя центромерами. Двойной сестринский обмен может привести к образованию двух сцепленных колец. В следующей интерфазе двойное кольцо может снова претерпеть один, два или более сестринских обмена, что в свою очередь приведет к двойным сцепленным кольцам или к четверным кольцам. Таким образом, возможно большое число разных вариантов. На рис. 2.53 представлено двойное сцепленное кольцо, на рис. 2.54, А - четверное кольцо. Многие из этих заново формирующихся колец ведут к нарушениям в митозе вследствие все большего числа разрывов и последующей анеуплоидии в дочерних клетках. На рис. 2.55 показана анафаза с разрывами дицентрических колец на равные и неравные части. Большая часть теоретически возможных конфигураций (рис. 2.40) действительно наблюдалась в данном случае. Фрагменты. Хромосомные фрагменты, не содержащие центромеры или ее части (так называемые ацентрические фрагменты), в митозе и мейозе обычно теряются, но при наличии центромеры они могут сегрегировать как дополнительные, маркерные, хромосомы. При исследовании случайной выборки новорожденных в Дании (разд. 5.1.2.1) такие маркеры оказались не редкими; в некоторых случаях у носителей этих маркерных хромосом обнаруживаются фенотипические аномалии. Изохромосомы. Иногда выявляются хромосомы, оба плеча которых идентичны. Их называют изохромосомами. Можно предположить, что они возникают вследствие аномального разделения метафазной хромосомы, как показано на рис. 2.56. Если в 86 2. Хромосомы человека
такую перестройку вовлекается неравноплечая хромосома, то может образоваться изохромосома и по короткому, и по длинному плечу. Относительно часто наблюдаются изохромосомы X. В случае изохромосомы по длинному плечу X, i(Xq) развивается синдром Тернера, поскольку данная хромосома всегда инактивирована и активной остается только одна нормальная Х-хромосома (разд. 2.2.3). Межхромосомные обмены: центрические слияния (робертсоновские транслокации). Центрическое слияние является наиболее частым типом хромосомных перестроек в человеческих популяциях. Первые описанные случаи транслокационного синдрома Дауна были связаны с центрическим слиянием между длинным плечом хромосомы 21 и одной из D- или G-хромосом. Впоследствии о таких больных сообщалось неоднократно. Среди всех случаев синдрома Дауна транслокации этого типа составляют всего лишь несколько процентов, и многие из них являются вновь возникшими. Важно, что в центрическое слияние Таблица 2.6. Хромосомы, вовлеченные в робертсоновские транслокации. (Семейный материал, проанализированный Шефером [501а].)
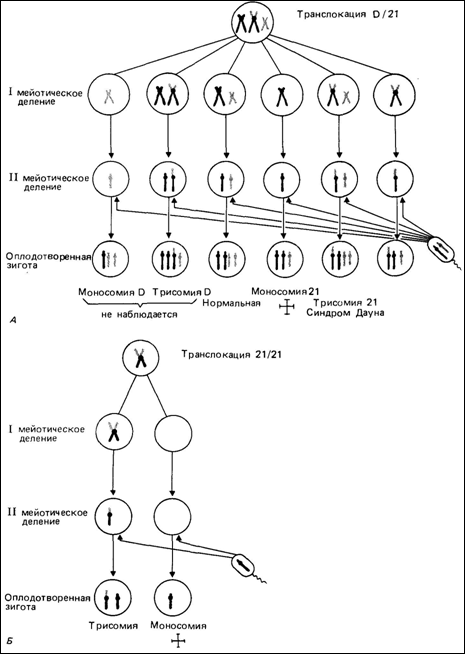
могут вовлекаться все пять пар акроцентрических хромосом. Короткие плечи этих хромосом содержат ядрышковые организаторы, в частности гены рРНК (разд. 2.3). При этом в интерфазном ядре короткие плечи, включая центромерные районы, располагаются в тесной близости от ядрышка. Благодаря применению методов дифференциального окрашивания появилась возможность исследовать участие отдельных D- и G-хромосом в центрических слияниях. Оказалось, что оно не является случайным (табл. 2.6). Данные, представленные в этой таблице, основаны на исследовании новорожденных. Следует учесть, что результаты могут быть искажены из-за неодинаковой частоты эмбриональной смертности в различных группах. Центрическое слияние означает, что короткие плечи двух акроцентрических хромосом и, вероятно, одна из центромер утрачены (рис. 2.57), т.е. утрачены также и гены рибосомной РНК. Действительно, по данным ДНК—РНК-гибридизации среднее число генов рРНК меньше у так называемых сбалансированных носителей центрических слияний, чем в общей популяции [1020, 1061]. Однако это не приводит к каким-либо функциональным различиям, и носители таких хромосом совершенно здоровы. На рис. 2.58 показаны возможные комбинации хромосом в половых клетках носителя D/Gи G21/G21-транслокации. После оплодотворения нормальным сперматозоидом возможны шесть разных вариантов. Однако первые два - моносомия D и трисомия D- никогда не наблюда- 2. Хромосомы человека 87
88 2. Хромосомы человека лись, а моносомия 21 по крайней мере в большей части известных случаев детальна. Каждый из остальных трех вариантов - трисомия 21, сбалансированная транслокация и нормальный набор- ожидается с вероятностью 1/3. Это ожидание, однако, не подтверждается на практике: когда мать является носителем, вероятность составляет около 15%, а если носитель отец, вероятность не превышает 5%. Однако риск появления сбалансированной транслокации составляет, как и ожидается, ~ 50%. При транслокации 21/21 (как и в случае 21/21 изохромосомы) прогнозы намного более мрачные: либо ребенок будет трисомиком с синдромом Дауна, либо анеуплоидия будет летальной. К счастью, в настоящее время можно при помощи методов дифференциального окрашивания отличить транслокацию 21/21 от транслокации 21/22, при которой вероятность анеуплоидных зигот намного меньше - такая же, как и в случае D/G-транслокаций. Межхромосомные обмены: реципрокные транслокации. В отличие от центрических слияний реципрокные транслокации не обязательно связаны с утратой материала. Фрагменты хромосом воссоединяются в новых комбинациях, но с сохранением в зиготе эуплоидного числа 46, а не 45, как при центрических слияниях. На рис. 2.60 представлены типы дочерних клеток, которые можно ожидать в случае реципрокных транслокаций. Чаще всего выявляются только частичные трисомики и частичные моносомики. Другие комбинации, как полагают, летальны. Типичный случай описан в работе [504]. На рис. 2.61 показаны два умственно отсталых сибса в возрасте 11 и 9 лет. В их фенотипе обнаружены как конкордантные, так и дискордантные признаки (табл. 2.7). При исследовании кариотипа обычным методом у обоих детей выявлено удлинение длинного плеча одной из С-хромосом (рис. 2.62); у матери и бабки (по линии матери) обнаружена такая же хромосома и, кроме того, другая аномальная хромосома в группе С (6-Х-12), у которой почти полностью отсутствовало короткое плечо (рис. 2.63). С помощью G-метода у матери выявлена реципрокная транслокация между хромосомами 7 и 10, кариотип 46, XX, t (7; 10) (р22; p11). Результатом такой перестройки является частичная трисомия Юр + у обоих детей. Особенность данного случая заключается не только в конкордантности многих признаков у обоих детей, что укладывается в единый клинический синдром, но и в наличии ряда дискордантных симптомов, что указывает на изменчивость фенотипических аномалий, вызванных одной и той же хромосомной аберрацией.
Основные фенотипические проявления аутосомных аберраций. Наиболее заметной особенностью фенотипов при аутосомных аберрациях является очень частое совпадение многих признаков и симптомов. Основные признаки: а) Общие низкий вес при рождении; резкая задержка развития; умственная отсталость (обычно тяжелая); низкий рост б) Голова и лицо микроцефалия; неполная оссификация; микрогнатия; аномальное расположение глаз; «дизморфическое лицо» 2. Хромосомы человека 89
90 2. Хромосомы человека
низко расположенные и деформированные ушные раковины в) Верхние и нижние конечности аномальный дерматоглифический рисунок г) Внутренние органы врожденный порок сердца и/или крупных сосудов пороки развития мозга пороки развития мочеполовой системы.
Следующие признаки обычно не указываются как характерные для аутосомных аномалий и описываются как исключения: умственная отсталость без каких-либо пороков развития пороки развития при нормальном психическом развитии изолированные (одиночные) пороки развития. При многих, хотя и не при всех аутосомных аберрациях, кроме этих общих пороков развития находят более или менее специфичные. Важно отметить, что и общие признаки могут проявляться с большей или меньшей тяжестью. Ряд признаков, причиной которых является специфическая аберрация, обычно формируют неслучайное сочетание, характерное именно для данной аберрации. Подозрение на хромосомный дефект может возникнуть при клиническом обследовании, но ставится диагноз только на основе хромосомного анализа. Наличие характерных симптомов указывает на необходимость исследования хромосом. У разных больных с одинаковой аберрацией многие характеристики одного синдрома сильно варьируют. Так, при синдроме Дауна, например, в ряде случаев умственная отсталость может быть выражена в незначительной степени, в то время как в большинстве случаев наблюдается олигофрения тяжелой степени; кроме того, пороки сердца находят у многих таких больных, а атрезию кишечника - крайне редко. У сибсов на рис. 2.61 с одной и той же транслокацией кроме некоторых общих черт обнаруживаются и явные фенотипические различия. Можно предположить, что эта изменчивость зависит частично от того, что у разных индивидов одна и та же аномальная хромосома проявляет свои эффекты на разном генетическом фоне. Наиболее неожиданный факт, касающийся фенотипа хромосомных аберраций, состоит в том, что при трисомиях вообще обнаруживаются аномалии. Ведь носители этих аберраций имеют полный набор генетического материала, и ни один из генов не утерян и не поврежден! По данным исследований гетерозигот по аутосомным
2. Хромосомы человека 91
92 2. Хромосомы человека рецессивным болезням (разд. 4.2.2.8) известно, что снижение активности ферментов вдвое, как правило, еще не нарушает их функции. В связи с этим трудно понять, почему увеличение количества генных продуктов в 1,5 раза, наблюдающееся при трисомиях, должно приводить к таким большим фенотипическим различиям. Более детально эти проблемы будут обсуждаться в разд. 4.7.4. Здесь только скажем, что симптомы, общие для всех аутосомных синдромов, не зависят от того, какая хромосома вовлечена в перестройку. Можно, впрочем, отметить, что пораженными оказываются чаще всего те системы органов, для которых характерен длинный и сложный путь эмбрионального развития, и, следовательно, для нормального обеспечения этого процесса необходимо много разных генов. Однако такое объяснение носит слишком общий характер, и к тому же все необходимые гены имеются в наличии. Чем же можно объяснить нарушения, вызываемые хромосомными аберрациями? Ответ таков: эти синдромы обусловлены, вероятно, не наличием избыточной активности или дефекта отдельных генов, а главным образом нарушениями регуляции активности генов во время эмбрионального развития. Следовательно, анализ аутосомных аберраций может оказаться полезным для понимания механизмов генной регуляции у человека. В случае такой специальной проблемы, как развитие половых признаков, изучение больных с численными и структурными аберрациями половых хромосом оказалось весьма поучительным. Однако до подробного обсуждения аберраций этого типа полезно сделать несколько замечаний относительно сегрегации и пренатальной селекции несбалансированных транслокаций, а также относительно возможных клинических признаков «сбалансированных» транслокаций. Кроме теоретического интереса эти вопросы важны с точки зрения генетического консультирования - для оценки повторного риска. Дата добавления: 2015-12-16 | Просмотры: 1770 | Нарушение авторских прав |