|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Частота мутаций и возраст отцаБлестящая идея Вайнберга. В одной из своих работ под названием "Zur Vererbung des Zwergwuchses" (О наследуемой карликовости - нем.) Вайнберг (1912) [1692] обсуждает генетические основы ахондроплазии. Материалы, которыми он располагал,это родословные, опубликованные Ришбитом и Баррингтоном (1912) [1599]. Вайнберг проверял предположение о простом рецессивном типе наследования ахондроплазии и в результате отказался от этой гипотезы. Он пришел к выводу, что эти данные лучше согласуются с предложением о дигибридном рецессивном наследовании. Вайнберг упомянул и о точке зрения Плэйта, который считал, что ахондроплазия наследуется по доминантному типу. Анализ имеющихся материалов показал, что более поздно родившиеся сибсы (родные братья и сестры - Прим. перев.) заболевают ею с большей вероятностью. Сделав несколько замечаний о возможных смещениях, он продолжал: «Если бы более точный анализ с учетом порядка рождения действительно подтвердил вывод о высокой частоте ахондроплазии у последних детей в семье, это указывало бы на то, что карликовость возникает в результате мутации» Примерно 30 лет спустя это предположение нашло подтверждение в работе Мерха [795], который проанализировал все случаи ахондроплазии в Дании, включая сведения о нескольких карликах, умерших за несколько лет до исследований. Он представил убедительные данные, свидетельствующие о том, что спорадические случаи этой патологии действительно возникли вследствие мутации de novo. Он также показал, что в этих спорадических случаях средний материнский, равно как и отцовский, возраст значительно выше популяционной средней и что эффект материнского 5. Мутации 171
возраста не связан с влиянием порядка рождения, который коррелирует как с отцовским, так и с материнским возрастом. Ему не удалось установить, обусловлен ли этот эффект возрастом матерей, отцов или и тех и других. Модель Уотсона — Крика стимулировала новые исследования влияния отцовского и материнского возраста. В 1953 г. Уотсон и Крик [1347] предложили свою модель структуры ДНК. Эта модель не только объясняет процессы репликации и хранения наследственной информации, но и позволяет выдвинуть убедительную гипотезу о механизме спонтанных мутаций, связывающую их возникновение с включением некомплементарных оснований при репликации. Такой механизм предполагает зависимость мутационного процесса от репликации. И действительно в ряде работ на микроорганизмах подтвердилось, что почти все мутации происходят в делящихся клетках [1668]. Эта концепция дала новый импульс статистическим исследованиям, посвященным изучению влияния отцовского возраста на мутационный процесс у человека. Касаясь данного вопроса, Пенроуз [1590а] высказал следующее соображение: «В женском зародышевом пути происходит очень мало клеточных делений, а в мужском - много, так как сперматогонии постоянно делятся. Поэтому частота мутаций, обусловленных неточным копированием гена в процессе клеточного деления, вряд ли испытывает скольконибудь сильное влияние материнского возраста; однако у отцов старших возрастов должно наблюдаться заметное увеличение числа дефектов, возникших по этой причине». Следствия из этой гипотезы о механизме мутаций можно сравнить с выводами, вытекающими из других возможных предположений о механизме их возникновения [1590а, 1668]. В табл. 5.9 каждой из пяти возможных гипотез поставлены в соответствие те или иные экспериментально-генетические результаты. Пенроуз рассматривал главным образом вторую модель (в которой мутации зависят от деления клеток). Она предсказывает увеличение частот мутаций с возрастом, происходящее только у мужчин, и более высокую мутационную
172 5. Мутации частоту в половых клетках мужчин, чем в половых клетках женщин. Клеточные деления в процессе развития половых клеток у мужчин и женщин. Для того чтобы более точно предсказать частоту мутаций, помимо правильного, но очень общего тезиса Пенроуза, мы должны знать число клеток и клеточных делений в зародышевых путях мужчин и женщин. В настоящее время данные из различных областей биологии и медицины, касающиеся этого вопроса, собраны воедино. По ним можно составить следующую картину ранней стадии развития половых клеток, оогенеза и сперматогенеза [1683]. Ранняя стадия развития. Первичные половые клетки человека выходят из желточного мешка на 27-й день после оплодотворения и мигрируют в гонадные складки. На 46-й день беременности гонады претерпевают половую дифференцировку и становятся либо яичниками, либо семенниками. Оогенез. Оогенез (рис. 5.15) протекает в течение внутриутробной жизни плода и прекращается к моменту рождения. После половой дифференцировки в результате митотических делений происходит быстрое увеличение числа стволовых клеток яичников. Начиная со 2-го месяца беременности то или иное число ооцитов вступает в профазу мейоза; оогонии, существующие свыше семи месяцев, претерпевают дегенерацию. Стадии лептотены и зиготены (см. разд. 2.1.2.4) приходятся на период между 2-м и 7-м месяцами беременности. После рождения все стволовые клетки, как правило, утилизируются, оогонии превращаются в ооциты или дегенерируют. Общая популяция половых клеток эмбриона увеличивается от 6 х 105 на 2-м месяце беременности до максимальной численности в 6,8 х 106 клеток во время 5-го месяца. Затем, к моменту рождения, эта популяция уменьшается до 2 х 106. Если принять правдоподобное предположение о пролиферации путем дихотомических делений, то можно рассчитать, что к моменту рождения оогонии разделится 22 раза. Расчет этого числа сводится к вычислению показателя степени, в которую надо возвести 2, чтобы получить приблизительную величину общей численности ооцитов, то есть 222 и 6,8 х 106. От рождения до половой зрелости и оплодотворения женская половая клетка претерпевает только два мейотических деления независимо от того, в каком возрасте происходит оплодотворение. Сперматогенез. При сперматогенезе (рис. 5.16) кинетика клеточных делений выглядит по-другому. На той стадии эмбрионального развития, на которой у женщин первичные половые клетки превращаются в оогонии, у мужчин они становятся гоноцитами. С ранней стадии эмбрионального развития и до половой зрелости семенные канальцы непрерывно пополняются так называемыми Ad-сперматогониями (от англ. d-darkтемный) и примерно к 16 годам происходит полное установление процесса сперматогенеза Число Ad-сперматогониев можно оценить тремя различными способами: по результатам измерения соответствующих объемов; на основании их
5 Мутации 173
среднего числа, приходящегося на поперечный срез канальца, и данных о длине канальца; а также из максимального числа сперматозоидов, продуцируемых в течение дня Величины этих оценок заключены в интервале от 4,3 х 108 до 6,4 х 108 в расчете на семенник. Приближенная оценка для обоих семенников составляет ≈ 1,2 х 109. Такое число клеток может образоваться в результате примерно 30 делений. Однако в отличие от ооцитов эти Ad-сперматогонии проходят непрерывную цепь делений Из двух продуктов деления одна клетка готовится к следующему делению на две Ad-клетки, тогда как другая делится, давая две Ар-клетки (от англ. p-pale - бледный). Последние развиваются в В-сперматогонии и сперматоциты, которые затем претерпевают мейотические деления (см. рис 5 16). Временные характеристики этих клеточных делений хорошо известны отчасти благодаря исследованиям m vivo, объектом которых служили молодые мужчины Цикл деления Ad-сперматогониев длится примерно 16 дней. Отсюда можно оценить число клеточных делений, соответствующее определенному возрасту (табл. 5.10). Если такой расчет дает приблизительно правильные результаты, то число делений сперматозоида от стадии раннего эмбрионального развития до 28 лет примерно в 15 раз больше числа делений, происходящих в течение цикла развития ооцита. Для мужчин старше данного возраста этот метод расчета даст еще более высокие величины. Такая экстраполяция довольно рискованна,
174 5. Мутации
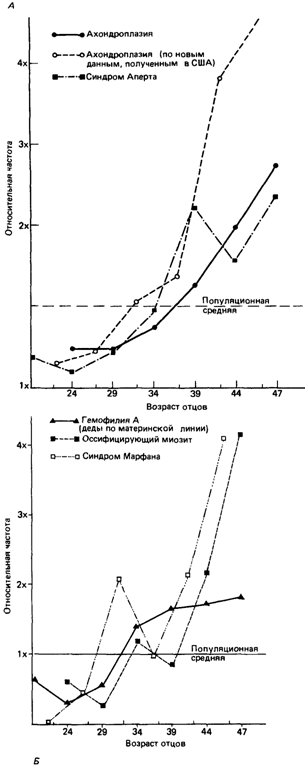
так как все исследования, на результатах которых основаны оценки, проводились на более молодых людях. Однако хорошо известно, что сексуальная активность, измеряемая числом эякуляций, понижается уже на 4-м десятилетии жизни. Некоторые данные свидетельствуют о том, что число сперматозоидов, подсчитанное в этом возрасте, по-видимому, немного увеличивается, но в целом, как показано на рис. 5.17, интенсивность сперматогенеза с возрастом несомненно понижается. Рис. 5.17 может служить также иллюстрацией кумулятивного распределения числа сперматогониальных делений, если замедление сперматогенеза обусловлено пролонгацией циклов делений Ad-сперматогониев. Возможны, однако, и другие механизмы такого замедления: например, может быть так, что одни Ad-сперматогонии дегенерируют, а другие продолжают делиться с прежней скоростью. Удивительно, но изменения процесса сперматогенеза, происходящие при старении мужчин, никогда не изучались с применением гистологических методов 1). Рост частоты мутаций с возрастом отца. Гистологические исследования необходимы, чтобы понять механизм, лежащий в основе реально наблюдаемого увеличения частоты мутаций с увеличением возраста отцов. На графиках, изображенных на рис. 5.18, нанесены относительные частоты мутаций, приведенные к соответствующей популяционной средней, для разных возрастных групп мужчин и нескольких болезней; помимо ахондроплазии, в их числе такие заболевания, как акроцефалосиндактилия (синдром Аперта) [1394], синдром Марфана [1570] и оссифицирующий миозит (болезнь, приводящая к прогрессирующему окостенению мышц) [1662] (см. табл. 5.11). Особенно интересен рост частоты спорадических случаев гемофилии А с увеличением возраста дедов по материнской линии [1486, 1598]. При проведении всех этих исследований возникали некоторые проблемы, связанные с получением адекватных контрольных выборок из общей популяции, так как большая часть популяционных статистических данных содержит информацию о числе новорожденных в зависимости от возраста матерей, а не отцов. Однако рассматриваемый эффект выражен столь отчетливо, что некоторое несоответствие контроля не окажет слишком сильного влияния на получаемый результат. Все кривые, изображенные на рис. 5.18, имеют две общие особенности: 1) частота мутаций в самой старшей груп- 1) Единственное исключение составляет знаменитая работа Фон Винивартера (1912) о числе хромосом человека [543], в которой изучался сперматогенез у четырех мужчин в возрасте 21, 23, 25 и 41 года. У первых трех он обнаружил все стадии процесса на всех сделанных им поперечных срезах семенных канальцев. Совершенно другая картина наблюдалась в случае 41-летнего мужчины. В то время как в некоторых участках сперматогенез протекал нормально, в других были найдены только сперматогонии и сперматоциты, а на срезах третьих вообще ничего не обнаружено, даже сперматогониев. Согласно имеющемуся описанию, этот тщательно изучавшийся пробанд был совершенно здоров и вел нормальную половую жизнь. Фон Винивартер не смог обнаружить в его семенниках каких-либо следов инфекции. Он сделал вывод, что эти изменения отражают нормальный процесс старения. Если этот единичный результат подтвердится, то характерный наклон кривой зависимости вероятности возникновения мутаций от возраста можно будет считать свидетельством в пользу гипотезы ошибок копирования.
176 5. Мутации
пе в несколько раз (приблизительно в пять) выше, чем в наиболее молодой группе; 2) наклон кривой роста проявляет тенденцию к прогрессирующему увеличению крутизны с увеличением возраста. Первая особенность согласуется с предположением, что этот рост обусловлен накоплением клеточных делений. Однако вторая особенность противоречит этому предположению; скорее следовало бы ожидать, что кривая выйдет на плато (по крайней мере в том случае, если скорость делений Ad-сперматогониев действительно замедляется с возрастом). Данное противоречие пока не нашло своего разрешения. Другие доминантные мутации, для которых возможен эффект отцовского возраста. Отцовский возраст может также служить ключом при идентификации доминантных мутаций, являясь фактором, способствующим возникновению синдромов, которые сопряжены с пороками развития и приводят к бездетности их носителей. К возможным кандидатам на роль доминантных патологий относятся гидроцефалии, за исключением расщелины позвоночника и микрофтальмии/анофтальмии [1590а]. К числу доминантных или возможно доминантных болезней, для которых предполагается существование эффекта отцовского возраста, принадлежат синдром базальноклеточного невуса, синдром Ваарденбурга, болезнь Кроузона, окулоденто-дигитальный синдром и синдром Тречера- Коллинза [1506].
Мутации, приводящие к возникновению неустойчивых гемоглобинов или гемоглобина М, и возраст отцов [1643]. Как отмечалось в разд. 4 3.2, гемоглобин М (метгемоглобин) и нестабильные гемоглобины служат причиной клинических синдромов, передающихся по аутосомно-доминантному типу. Стаматояннопулос собрал обширную информацию о родословных, анализ которых показывает, что один из этих гемоглобинов появился в результате мутации de novo. В общей сложности были собраны данные о 50 проживающих в 14 странах индивидах, имеющих такие гемоглобины Все они родились между 1922 и 1976 гг. Средний возраст отцов 32,7 года, а матерей - 28,3 года. Для того чтобы сравнить возраст родителей пробандов с возрастом родителей из общей контрольной популяции, авторы рассчитали для каждого года и каждой страны кумулятивные частотные распределения возраста всех родителей. Возраст отца и матери каждого пробанда выражали в процентилях этих распределений. Процентили распределения отцовских возрастов смещены к верхнему концу интервала, 11 из 50 возрастных групп отцов пробандов заключены между 90-м и 100-м процентилями (рис. 5.19). Хотя этот результат наводил на мысль о существовании эффекта отцовского возраста, отличие данного распределения от контрольного не достоверно. Адекватная проверка отцовства в большинстве случаев невозможна. Следовательно, среди этих пробандов могло оказаться заметное число «ложных мутантов», появившихся вследствие ошибочных предположений об отцовстве. Позднее были опубликованы данные о новой мутации одного из Нb М; отцу мутантного ребенка было 49 лет, а матери - 37 [1586]. Опубликованы чрезвычайно подробно документированные сообщения о мутации de novo в случае β-талассемии [1657; 1658] Клинические и биохимические обследования ребенка, которому к моменту постановки диагноза 5. Мутации 177
исполнилось 2 года, а также обоих родителей и трех сибсов не оставили никаких обоснованных сомнений в том, что это действительно новая мутация; ложное отцовство полностью исключено. Ко времени рождения пробанда отцу было 45 лет, а матери - 44 года. В том же 1965 году в Швейцарии средний возраст отцов был равен 31 году, а матерей - 28,2. Сообщение о единичном случае не может заменить статистического исследования; тем не менее этот результат - убедительное свидетельство в пользу существования эффекта отцовского возраста. Однако знание того, что мутации de novo с большей вероятностью возникают у отцов старших возрастов, может привести к перекосу в сторону публикации сведений о таких отдельных случаях.
Некоторые доминантные мутации обнаруживают слабый эффект отцовского возраста. Пенроуз [1590] отметил, что не все доминантные мутации проявляют сильный эффект отцовского возраста. К заболеваниям, для которых этот эффект выражен гораздо слабее, относится такая хорошо изученная болезнь, как двусторонняя ретинобластома [1683]. В числе других-туберозный склероз, нейрофиброматоз и несовершенный остеогенез. На рис. 5.20 приведены данные о соответствующих частотах мутаций. Для трех последних из вышеупомянутых болезней увеличение частот с возрастом отцов не достоверно; однако обнаружен достоверный эффект порядка рождения, свидетельствующий все-таки в пользу гипотезы о влиянии отцовского возраста. Сравнение рисунков 5.18 и 5.20 приводит к обоснованному выводу о гетерогенности доминантных мутаций в отношении эффекта отцовского возраста. Вероятно, есть разные типы мутаций: одни обнаруживают сильный отцовский эффект, а другие - слабый или вообще его не проявляют. Из факта существования эффекта отцовского возраста для летальных или полулетальных мутаций Х-сцепленных генов следуют важные выводы. Если предположить, что в поколении дедов мутации происходили чаще, чем в поколении отцов, можно ожидать, что эффект отцовского возраста будет часто обнаруживаться на дедах пробандов по материнской линии.
178 5. Мутации Кроме того, если мутации зависят от репликации, то они должны гораздо чаще встречаться у дедов, чем у бабок, так как при сперматогенезе происходит намного больше клеточных делений, чем при оогенезе. Эта проблема изучалась с использованием двух групп данных о 77 индивидах с гемофилией А, среди которых предполагалось найти мутанта de novo [1486; 1598]. С увеличением возраста дедов действительно наблюдался существенный рост частоты мутаций (рис. 5.18, Б), что нашло подтверждение в одной из последующих работ. Это еще один аргумент в пользу предположения о более высокой частоте мутаций в половых клетках мужчин (см. ниже). Другая Х-сцепленная патология: синдром Леша — Напхана (30800). Синдром Леша—Найхана - болезнь, сцепленная с Х-хромосомой, - возникает в результате дефекта гипоксантин-гуанин—фосфорибозилтрансферазы. Этот дефект можно диагностировать не только у мужчин, являющихся, как известно, гемизиготами, но и в клетках гетерозиготных женщин с лайонизированной Х-хромосомой. В рассматриваемом ниже исследовании [1456] обнаружено пять случаев, когда мать пробанда была гетерозиготой, а бабка по материнской линии - нормальной гомозиготой. Следовательно, мутация должна была возникнуть в половой клетке деда со стороны матери. Возраст дедов и бабок
приведен в табл. 5.12. Соответствующие средние величины гораздо выше усредненных значений для популяции Соединенных Штатов. Таким образом, весьма вероятно, что для синдрома Леша—Найхана величина эффекта отцовского возраста имеет такой же порядок, как и в случае других рассмотренных выше болезней. Дата добавления: 2015-12-16 | Просмотры: 808 | Нарушение авторских прав |

 )-атрофированные клетки.
)-атрофированные клетки.