|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Определение поляРазвитие половых признаков. Формирование половых признаков у человека – один из аспектов его развития, изученный относительно хорошо. Эта область представляет собой пример удачного сочетания теории, наблюдений непосредственно на человеке и экспериментов на животных. Теория подсказывала исследователям направления экспериментов, которые в свою очередь позволяли усовершенствовать саму теорию. Наблюдения генетических аномалий определения пола у человека тоже способствовали развитию теории и, кроме того, указывали на необходимость новых экспериментов. Принято различать четыре уровня половой дифференцировки: 1) хромосомное определение пола (46 XX или 46 XY); 2) определение пола на уровне гонад (яичники или семенники); 3) фенотипическое определение пола (женский или мужской; внешние половые признаки); 4) психологическое определение пола. Четвертый уровень будет обсуждаться в разд. 8.2.2.3, первый частично уже затрагивался в разд. 2.2.3. Как отмечалось, анализ аутосомных аберраций, несмотря на его несомненную важность для понимания механизмов нормального развития и его нарушений, до сих пор не позволил выяснить ничего конкретного о механизмах развития. С другой стороны, анализ нарушения числа и структуры половых хромосом дал очень много ценной информации не только о хромосомном определении пола (1-й уровень), но также об определении пола на уровне гонад и фенотипа (уровни 2 и 3, см. ниже и [1068а]). Зачатки гонад у ранних эмбрионов (до 5-й или 6-й недели) не различаются у разных полов и не содержат клеток зародышевого пути. Первичные клетки зародышевого пути у человека можно обнаружить на 3-й неделе эмбрионального развития в эктодерме желточного мешка. Затем под влиянием хемотаксических сигналов они мигрируют в гонады. Эта миграция не зависит от пола; в соответствующих экспериментальных системах женские половые клетки мигрируют в мужские гонады и наоборот. Зачатки гонад могут развиваться в яичники или семенники (уровень 2). В норме направление развития определяется наличием Y-хромосомы; мужские гонады развиваются, если имеется одна Y-хромосома, независимо от числа Х-хромосом. Согласно современным данным, это развитие зависит от H-Y-антигена. Y-хромосома и H-Y-антиген. Эйчвальд и Силмсер в 1955 г. описали у мыши трансплантационный антиген, контролируемый Y-хромосомой (разд. 3.1.4) [643]. Лишь значительно позднее стало понятным его значение для определения пола и особенно для дифференцировки мужских гонад [1248; 1249; 1341; 1343; 1342]. В настоящее время H-Y-антиген, определяющий пол, обнаружен у нескольких видов. По-видимому, его секретируют мужские первичные клетки зародышевого пути. Как только эти клетки попадают в зачатки гонад, начинается дифференцировка семенников. Этот вывод в значительной степени основан на изучении случаев отклонений в развитии пола, например у фримартин (т.е. коров генетически женского пола, у которых гонады превратились в семенники в результате того, что некоторое количество клеток близнеца мужского пола транспортировалось через общую кровеносную систему в гонады женского эмбриона). В настоящее время получены экспериментальные данные в пользу того, что H-Y-рецепторы имеются на поверхности клеток гонад обоих типов. Совместная инкубация дезинтегрированной ткани семенников с H-Y-антигеном приводит к сборке структур, напоминающих семенники. Такая сборка может быть индуцирована и в случае клеток женских гонад. Если активность H-Y-антигена подавить добавлением анти-Н-Y-антисыворотки, то возникают структуры, характерные для яичников. Данные, полученные в последнее время для редких случаев в определении пола (например, для мужчин с генотипом XX), удается объяснить, предположив, что H-Y-антиген кодируется не Y-хромосомой, а структурным аутосомным геном, который находится под контролем Y-хромосомы. Этот ген может быть репрессирован у всех индивидов, не имеющих Y-хромосомы; его экспрессия может быть индуцирована факторами, которые в норме определяются Y-хромосомой. Следователь- 138 4. Действие генов
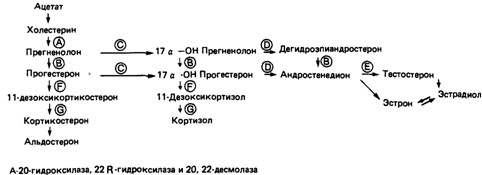
но, возможны конститутивные мутации, при которых будет синтезироваться H-Y-антиген, приводя к развитию мужского фенотипа даже в тех случаях, когда все клетки организма лишены Y-хромосомы. Возможно, однако, что произошла транслокация, в результате которой ген H-Y-антигена переместился с Y-хромосомы на аутосому. С другой стороны, H-Y-антиген обнаруживали (хотя и в низком титре) даже у женщин с синдромом Тернера (ХО), а также у XX-мужчин. Эти и другие наблюдения привели к выводу, что определение пола на уровне гонад может быть пороговым феноменом; для превращения зачатка гонад в семенники необходима некоторая минимальная концентрация H-Y-антигена. В последнее время роль H-Y-антигена в формировании семенников вызывает некоторые сомнения, которые связаны с новыми наблюдениями на животных и человеке. Например, H-Y-антигены, обусловливающие реакцию отторжения трансплантата и вызывающие цитотоксический эффект, вероятно, не идентичны, поскольку у некоторых животных эти два теста дают противоположные результаты [1114]. Следует учесть, однако, что тест на цитотоксичность связан с практическими трудностями и несколько субъективен [1038]. Развитие вторичных половых признаков. Развитие половых признаков обусловлено дифференцировкой гонад. Половые органы формируются из мюллеровых и вольфовых протоков, которые происходят из первичной почки. У женщин мюллеровы протоки развиваются в фаллопиевы трубы и матку, а вольфовы протоки атрофируются. У мужчин вольфовы протоки развиваются в семенные протоки и семенные пузырьки. Под влиянием хорионического гонадотропина матери клетки Лейдига в эмбриональных семенниках синтезируют стероидные гормоны тестостерон и 5-дигидротестостерон. В клетках Сертоли синтезируется гормон, который называют мюллеровым ингибирующим фактором (MIF). Эти гормоны действуют на бипотентные зачатки внешних и внутренних половых органов, прежде всего на вольфовы протоки, мюллеровы протоки и мочеполовой синус. Нормальные индивиды мужского пола развиваются, только если все эти элементы функционируют в нужное время и в надлежащем месте. При их полном отсутствии формируются женские половые признаки; таким образом, развитие женских половых признаков не требует специальных регуляторных факторов, оно в этом смысле является «конститутивным». Незначительные отклонения в работе этой системы на различных уровнях вызывают неполное развитие мужского фенотипа в организме с мужским генотипом (мужской псевдогермафродитизм); анализ таких аномалий позволил получить обширную информацию о нормальной физиологии развития пола. Джост писал по этому поводу: «Становление мужского организма – это длительное, нелегкое и рискованное предприятие, своего рода борьба против имманентного стремления к женственности». Известно по меньшей мере 19 различных дефектов генов, аутосомно-рецессивных или сцепленных с Х-хромосомой, которые вызывают нарушения дифференцировки внешних и внутренних мужских половых признаков. Например, нарушенными могут быть и синтез андрогенов и синтез хорионического гонадотропина, могут отсутствовать рецепторы для этого гормона на клетках Лейдига; дефектными могут оказаться и пять ферментов, участвующих в синтезе тестостерона. Причиной аномалий может быть и нечувствительность клеток вольфовых протоков или мочеполового синуса к тестостерону или 5-дигидротестостерону вследствие дефектов рецепторов. Пути биосинтеза тестостерона и соответствующие генетические блоки приведены на рис. 4.73. В левой части рисунка (G, F) показаны нарушения, характерные для адреногенитальных синдромов у женщин, но не для псевдогермафродитизма у мужчин. Нарушения, обозначенные А (наследственная липоидная гиперплазия надпочечников), изучены мало; такие индивиды с мужским генотипом не только имеют женские наружные половые органы, но также страдают от сильных нарушений солевого обмена. Это справедливо и для блока В. В правой части рисунка показаны нарушения, которые вызывают псевдогермафро- 4. Действие генов 139
дитизм различной степени, не сопровождающийся другими проявлениями адреногенитального синдрома. Особого внимания заслуживает аутосомно-рецессивный признак - дефект 5-α-редуктазы (не показан на рис. 4.73). В норме этот фермент восстанавливает тестостерон до 5-α-дигидротестостерона в клетках мочеполового синуса. При его отсутствии развиваются нормальные мужские внутренние половые органы (семенные пузырьки, простата), а также вторичные мужские половые признаки, включая развитие мускулатуры, характер волосяного покрова и т.д., за исключением внешних половых органов, которые при поверхностном обследовании можно принять за женские. Отсюда название псевдовагинальная перинеоскротальная гипоспадия (26460). Эти дефекты ферментов встречаются редко. Существуют более частые синдромы, при которых андрогены не изменены, но соответствующие ткани-мишени полностью или частично невосприимчивы к ним. Синдром тестикулярной феминизации (31370) [1114]. При рождении эта аномалия никак не проявляется; больные выглядят как обычные девочки и в детском возрасте аномалию обычно удается идентифицировать, только если при паховых грыжах обнаруживаются семенники. Больные с таким синдромом имеют мужской кариотип, и мужские гонады. Термин «тестикулярная феминизация» был предложен Моррисом в 1953 г. [1220]. С наступлением половой зрелости отмечается аменорея, а в большинстве случаев обращает на себя внимание также полное или частичное отсутствие подмышечных волос, волос в области лобка и на теле. У взрослых рост и пропорции типично женские, хотя ноги часто несколько длиннее. Молочные железы хорошо развиты. Пропорции тела таких индивидов соответствуют скорее современным представлениям о женской красоте, чем среднему телосложению, поэтому неудивительно, что больные неоднократно встречались среди манекенщиц. Влагалище обычно укорочено и заканчивается слепым мешком. Вместо матки часто имеются остатки мюллеровых протоков, а вместо фаллопиевых труб можно найти мышечно-волокнистый тяж. Семенники локализуются в больших половых губах, паховом канале или в брюшной полости, и могут содержать нормальное или даже увеличенное количество клеток Лейдига, продуцирующих гормоны. Сперматогенез обычно отсутствует. Иногда наблюдаются злокачественные опухоли семенников. У больных андрогены секретируются в нормальных количествах, в особенности тестостерон, и потому можно было бы 140 4 Действие генов
ожидать нормального мужского развития. Наиболее очевидное объяснение механизма этой болезни, которое приводилось в течение многих лет, заключается в ненормальной конечной реакции органов на гормон. Недавно был обнаружен дефект рецепторов андрогенов [1359]. Это подтвердилось в различных экспериментах: например, лечение тестостероном не позволяет добиться изменений в голосе, росте бороды, гипертрофии клитора или (у больных мышей) индукции алкогольдегидрогеназы в почках. Психологическое развитие при тестикулярной феминизации происходит целиком по женскому типу. В настоящее время получены данные, указывающие, что взаимодействие андрогенов и их рецепторов влияет на развитие мозга, электроэнцефалограмму и поведение (разд. 8.2.2.3). Многие случаи таких отклонений встречаются чисто случайно, однако иногда наблюдаются семейные корреляции, при этом обнаруживается наследование от здоровых женщин и дополнительные случаи среди сестер матерей (матроклинные тетки пробандов). Это позволяло предположить мутацию, сцепленную с Х-хромосомой. Однако, поскольку репродукция у больных полностью нарушена, не удавалось получить и окончательных доказательств сцепления с Х-хромосомой, т.е. наблюдать наследование признака от гемизиготы всеми дочерьми, но не сыновьями. Поэтому нельзя исключить ограниченного полом проявления аутосомно-доминантного гена. Однако, в 1971 г. у мыши была обнаружена мутация с очень близким фенотипом [1025], которая явно была сцеплена с Х-хромосомой. Х-хромосома млекопитающих остается эволюционно стабильной, расположенные в ней гены являются гомологичными у всех до сих пор изученных видов млекопитающих [156]. Более того, рецепторы тестостерона можно обнаружить также в фибробластах, причем гетерозиготы имеют две разные популяции фибробластов - нормальную и неспособную связывать тестостерон, как это предсказывалось гипотезой Лайон [985]. Таким образом, сцепление с Х-хромосомой можно считать установленным. Любопытно, что Паттерсон и Бонньер, основываясь на анализе родословных, еще в 1937 г.
предполагали (тогда определение пола по генотипу было еще невозможно), что больные являются генотипически мужчинами и признак либо наследуется сцепленно с Х-хромосомой, либо ограничен полом. В результате новых мутаций могут появляться спорадические случаи, давление отбора в этом случае довольно значительное, больные бесплодны, поэтому можно ожидать обнаружения многочисленных новых мутаций (разд. 5.1.3). Генетическая гетерогенность Полипептидные гормоны, такие как инсулин, связываются с рецепторами, расположенными на мембранах клеток-мишеней (разд 3 8 14) С другой стороны, стероидные гормоны, как например, тестостерон, после проникновения в клетку путем диффузии связываются с цитоплазматическими рецепторами. На рис. 4.74 показан путь тестостерона и дигидротестерона в клетке, начиная со связывания с цитоплазматическим 8S-рецептором, через 4S-рецепторный комплекс к 5S- 4. Действие генов 141
рецепторному комплексу, который транспортируется в ядро. Большинство мутаций, приводящих к тестикулярной феминизации, затрагивают 8S-рецептор. Вполне вероятно, что мутантными могут быть 4S- и 5S-комплексы. Действительно, зарегистрированы случаи тестикулярной феминизации, при которых 8S-рецептор нормален. Кроме того, известны больные с неполной тестикулярной феминизацией, для которых характерно наличие интерсексуальных гениталий. При этом 8S-рецепторы, хотя и обнаруживаются, но в меньшем количестве. Мы коснулись лишь немногих генетических нарушений определения пола у мужчин. Известно много других, которые согласуются с теоретической концепцией становления пола и благодаря которым мы составили себе представления об этом процессе. Даже у фенотипически нормальных мужчин иногда встречаются генетические дефекты, вызывающие стерильность, например, нарушение формирования из половых клеток жизнеспособных сперматозоидов [1032].
Мутации
Дата добавления: 2015-12-16 | Просмотры: 690 | Нарушение авторских прав |