|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология
|
Полигенное наследования количественных признаков
- плейотропия
- экспрессивность и пенетрантность генов
Большинство количественных признаков организмов определяется несколькими неаллельнимы генами (полигенами). Взаимодействие таких генов в процессе формирования признака называется полимерным. В этом случае две или более доминантных аллели в равной степени влияют на развитие одной и того же признаки. Поэтому полимерные гены принято обозначать одной буквой латинского алфавита с цифровым индексом, например: А1А1 и а1а1. Впервые однозначные факторы были выявлены шведским генетиком Нильсон-Эле (1908 г.) при изучении наследования цвета в пшеницы. Было установлено, что этот признак зависит от двух полимерных генов, поэтому при скрещивании доминантних и рецессивных дигомозигот - окрашенной (А1А1, А2 А2) с бесцветной (а1а1, а2а2) - в F, все растения дают окрашенные семена, хотя они светлее, чем родительские экземпляры, которые имеют красное семя. В F, при скрещивании особей первого поколения проявляется расщепление по фенотипу в соотношении 15:1, потому бесцветным является лишь рецессивные дигомозиготы (а1а1 а2а2). В пигментированных экземплярах интенсивность цвета очень отличается в зависимости от количества полученных ими доминантних аллелей: максимальная в доминантных дигомозигот (А1А1, А2 А2) и минимальная у носителей одного из доминантных аллелей.
Важная особенность полимерии - суммация действия неаллельних генов на развитие количественных признаков. Если при моногенном наследовании признака возможны три варианта "доз" гена в генотипе: АА, Аа, аа, то при полигенных количество их возрастает до четырех и более. Суммация "доз" полимерных генов обесчивает существования непрерывных рядов количественных изменений.
Биологическое значение полимерии заключается еще и в том, что признаки, кодируемые этими генами, более стабильны, чем те, которые кодируются одним геном. Организм без полимерных генов был бы очень неустойчивым: любая мутация или рекомбинация приводила бы к резкой изменчивости, а это в большинстве случаев имеет неблагоприятный характер.
У животных и растений есть много полигенных признаков, среди них и ценные для хозяйства: интенсивность роста, скороспелость, яйценоскость, количество молока, содержание сахаристых веществ и витаминов и т.п.
Пигментация кожи у человека определяется пятью или шестью полимерными генами. В коренных жителей Африки (негроидной расы) преобладают доминантные аллели, у представителей европеоидной расы - рецессивные. Поэтому мулаты имеют промежуточную пигментацию, но при браках мулатов у них возможно появление как более, так и менее интенсивно пигментированных детей.
Многие морфологические, физиологические и патологические особенности человека определяются полимерными генами: рост, масса тела, величина артериального давления и др. Развитие таких признаков у человека подчиняется общим законам полигенного наследования и зависит от условий среды. В этих в случаях наблюдается, например, склонность к гипертонической болезни, ожирению и др. Данные признаки при благоприятных условиях среды могут не проявиться или проявиться незначительно. Эти полигенные признаки отличаются от моногенных. Изменяя условия среды можно обеспечить профилактику ряда полигенных заболеваний.
Плейотропия
Плейотропное действие генов - это зависимость нескольких признаков от одного гена, то есть множественное действие одного гена. В дрозофилы ген белого цвета глаз одновременно влияет на цвет тела, длины, крыльев, строение полового аппарата, снижает плодовитость, уменьшает продолжительность жизни. У человека известна наследственная болезнь - арахнодактилия ("паучьи пальцы"-очень тонкие и длинные пальцы), или болезнь Марфана. Ген, отвечающий за эту болезнь, вызывает нарушение развития соединительной ткани и одновременно влияет на развитие нескольких признаков: нарушение строения хрусталика глаза, аномалии в сердечно-сосудистой системе.
Плейотропное действие гена может быть первичным и вторичным. При первичной плейотропии ген проявляет свой множественный эффект. Например, при болезни Хартнупа мутация гена приводит к нарушению всасывания аминокислоты триптофана в кишечнике и его реабсорбции в почечных канальцах. При этом поражаются одновременно мембраны эпителиальных клеток кишечника и почечных канальцев с расстройствами пищеварительной и выделительной систем.
При вторичной плейотропии есть один первичный фенотипний проявление гена, вслед за которым развивается ступенчатый процесс вторичных изменений, приводящих к множественным эффектам. Так, при серповидно клеточной анемии у гомозигот наблюдается несколько патологических признаков: анемия, увеличенная селезенка, поражение кожи, сердца, почек и мозга. Поэтому гомозиготы с геном серповидно клеточной анемии гибнут, как правило, в детском возрасте. Все эти фенотипные проявления гена составляют иерархию вторичных проявлений. Первопричиной, непосредственным фенотипним проявлением дефектного гена является аномальный гемоглобин и эритроциты серповидной формы. Вследствие этого происходят последовательно другие патологические процессы: слипание и разрушение эритроцитов, анемия, дефекты в почках, сердце, мозге - эти патологические признаки вторичны.
При плейотропии, ген, воздействуя на какой то один основнй признак, может также менять, модифицировать проявление других генов, в связи с чем введено понятие о генах-модификаторах. Последние усиливают или ослабляют развитие признаков, кодируемых "основным" геном.
Показателями зависимости функционирования наследственных задатков от характеристик генотипа является пенетрантность и экспрессивность.
Рассматривая действие генов, их аллелей необходимо учитывать и модифицирующее влияние среды, в которой розвивается организм. Если растения примулы скрещивать при температуре 15-20 ° С, то в F1 согласно менделивской схеме, все поколения будут иметь розовые цветы. Но когда такое скрещивание проводить при температуре 35 °С, то все гибриды будут иметь цветы белого цвета. Если же осуществлять скрещивания при температуре около 30 ° С, то возникает разное соотношение (от 3:1 до 100%) растений с белыми цветами.
Такое колебание классов при расщеплении в зависимости от условий среды получило название пенетрантность - сила фенотипного проявления. Итак, пенетрантность - это частота проявления гена, явление появления или отсутствия признака у организмов, одинаковых по генотипу.
Пенетрантность значительно колеблется как среди доминантных, так и среди рецессивных генов. Наряду с генами, фенотип которых появляется только при сочетании определенных условий и достаточно редких внешних условий (высокая пенетрантность), у человека есть гены, фенотипное проявление которых происходит при любых соединениях внешних условий (низкая пенетрантность). Пенетрантностью измеряется процентом организмов с фенотипным признаком от общего количества обследованных носителей соответствующих аллелей.
Если ген полностью, независимо от окружающей среды, определяет фенотипное проявление, то он имеет пенетрантность 100 процентов. Однако некоторые доминантные гены проявляются менее регулярно. Так, полидактилия имеет четкое вертикальное наследования, но бывают пропуски поколений. Доминантная аномалия - преждевременное половое созревание - присуще только мужчинам, однако иногда может передаться заболевания от человека, который не страдал этой патологией. Пенетрантностью указывает, в каком проценте носителей гена оказывается соответствующий фенотип. Итак, пенетрантность зависит от генов, от среды, от того и другого. Таким образом, это не константное свойство гена, а функция генов в конкретных условиях среды.
Экспрессивность (лат. ехргеssio - выражение) - это изменение количественного проявления признака в разных особей-носителей соответствующего аллелей.
При доминантных наследственных заболеваниях экспрессивность может колебаться. В одной и той же рсемье могут проявляться наследственные болезни от легких, едва заметных до тяжелых: различные формы гипертонии, шизофрении, сахарного диабета и т.д. Рецессивные наследственные заболевания в пределах семьи проявляются однотипно и имеют незначительные колебанийния экспрессивности.
Билет 7
Генотип, фенотип. Закон расщепления:
Второй закон Менделя, заключающийся в том, что во второмпоколении гибридов при моногибридном скрещивании признак в случае промежуточного наследованияпроявляется в соотношении 1:2:1 (25% — гомозиготны как у одного родителя, 25 — как у другого и 50% —гетерозиготны), а в случае доминирования одного из аллелей — 1:3.
Влияние на расщ.по ф/типу – см. Прошлый билет.
Билет 8
Летальные гены. Изменение расщепления по фенотипу в отношении 3:1 во втором поколении моногибридного скрещивания бывает связано с разной жизнеспособностью зигот F2- Разная жизнеспособность зигот может быть обусловлена наличием ле-tsuibHbix генов. Летальным называется ген, вызывающий нарушения в развитии организма, что приводит его к гибели или уродству.
Изучение врожденных аномалий показало, что при разных летальных генах гибель особей бывает различной и может происходить на разных стадиях развития.
По классификации, предложенной Розенбауэром (1969), гены, вызывающие гибель 100 % особей до достижения ими половой зрелости, называются летальными, более 50 % — сублетальными (полулетальными) и менее 50 % — субвитальными. Однако следует отметить, что разделение это до некоторой степени условно и иногда не имеет четких границ. Примером может служить сцепленная с полом голость у кур. Почти половина голых цыплят гибнет в последние 2—3 дня инкубации. Из числа вылупившихся около половины цыплят гибнет до 6-недельного возраста, если их выращивают при температуре 32—35 "С. Но если температура в брудерах будет повышена на 5,5 "С, то погибнет значительно меньше голых цыплят. В 4—5 мес у голых цыплят вырастает редкое оперение и они уже в состоянии переносить довольно низкие температуры. В естественных условиях эта мутация, по-видимому, будет летальной и приведет к 100%-ной гибели птиц. Приведенный пример показывает, что характер проявления полулетального гена в значительной мере может зависеть от условий среды.
Летальные гены могут быть доминантными и рецессивными. В числе первых летальных факторов был открыт аллель, обусловливающий желтую окраску мышей. Ген желтой окраски является доминантным (Y). Его рецессивный аллель (у) в гомозиготном состоянии обусловливает появление черной окраски. Скрещивание желтых мышей между собой давало в потомстве две части особей желтых и одну часть черных, т. е. получалось расщепление 2:1, а не 3:1, как следовало из правила Менделя. Оказалось, что все взрослые мыши гетерозиготны (Yy). При скрещивании между собой они должны были дать одну часть гомозиготного потомства по желтой окраске (YY), но оно погибает еще в эмбриональном периоде, две части гетерозигот (Yy) будут желтыми и одна часть гомозигот по рецессивному признаку (уу) будет черной. Схема скрещивания выглядит так:
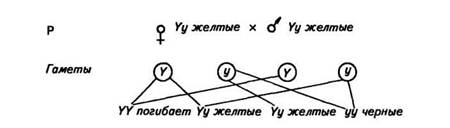
Таким же образом наследуется серая окраска шерсти у каракульских овец (сокольских, малич и др.), платиновая окраска у лисиц, распределение чешуи у линейных карпов и т. д.
Летальные гены в большинстве случаев рецессивны и поэтому долгое время могут находиться в скрытом состоянии. Совершенно здоровое и нормальное по фенотипу животное может быть носителем летального гена, действие которого обнаруживается только при переходе в гомозиготное состояние. В гомозиготное состояние летальные гены чаще всего переходят при родственном спаривании. В практике животноводства при разведении лошадей был случай гибели 25 жеребят на 2—4-й день после рождения от уродства прямой кишки — отсутствия заднепроходного отверстия (Atresia ani). Оказалось, что все жеребцы и кобылы, от которых рождались такие ненормальные жеребята, происходили от одного жеребца. Он был гетерозиготен по летальному гену (LI). Вначале этот жеребец при скрещивании с нормальными кобылами (LL) дал потомство, по фенотипу нормальное, но по генотипу половина потомков была благополучной (LL), a половина — гетерозиготной (LI), несущей рецессивный задаток (/) летального гена. При родственном спаривании гетерозиготных животных (Ы х Ы) появилась часть жеребят, гомозиготных по летальному гену (It), с уродством прямой кишки. Они все погибли. (Более подробно об аномалиях при летальных генах будет сказано в соответствующей главе.)
Билет 9.
Закон чистоты гамет: в каждую гамету попадает только одна аллель из пары аллелей данного гена родительской особи.
РЕЦИПРОКНОЕ СКРЕЩИВАНИЯ (от лат. reciprocus - взаимный), два скрещивания, которые характеризуются взаимно противоположным сочетанием анализируемого признака и пола (или типа спаривания) у форм, принимающих участие в этих скрещиваниях. Так, если в одном скрещивании у животных самка имела доминантный признак, а самец - рецессивный, то во втором скрещивании, реципрокном первому, самка должна иметь рецессивный признак, а самец - доминантный. Реципрокное скрещивание используют в генетическом анализе для выявления наследств, факторов, локализованных в Х-хромосоме. В этом случае в одном из реципрокного скрещивания Наблюдают явление«крисс-кросс» (крест-накрест) наследование, когда материнский признак передаётся только сыновьям, я отцовский - только дочерям. Кроме того, реципрокные скрещивания позволяют локализовать цитоплазматические наследственные факторы в случае анизогамии, когда в обоих реципрокных скрещиваниях наблюдают передачу потомкам только материнского признака.
Скрещивание гибридов первого поколения с особями, сходными по генотипу с родственными формами, называется ВОЗВРАТНЫМ.
P ♀AA × ♂aa
G A a
F1 Аа
Скрещивание потомков первого поколения с рецессивной родительской формой называется АНАЛИЗИРУЮЩИМ. Анализирующие скрещивание широко применяется при гибридологическом анализе, когда нужно установить генотип интересующей нас особи. Например, при необходимости определить, гомозиготным (АА) или гетерозиготным (Аа) является белый кролик, его скрещивают с черной крольчихой (аа). Если кролик гомозиготный, то все потомки будут белыми. Если он гетерозиготный, то появляются крольчата и белые и черные.
P ♂AA белые × ♀ aa черные ♂Aа белые × ♀ aa черные
G A a A a а
F1 Аа белые Аа белые аа черные
Билет 10
Дата добавления: 2015-12-16 | Просмотры: 892 | Нарушение авторских прав
|