|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
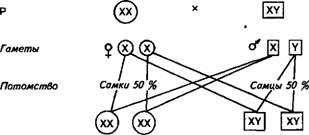
Дерминация пола и механизм его наследованияДЕТЕРМ ПОЛА: В ходе эволюции у большинства раздельнополых организмов сформировался механизм детерминации пола, обеспечивающий образование равного количества самцов и самок, что необходимо для нормального самовоспроизведения вида. Детерминация пола может происходить на разных этапах размножения. Схема образования гамет в мейозе самцов и самок, определения пола при оплодотворении следующая:
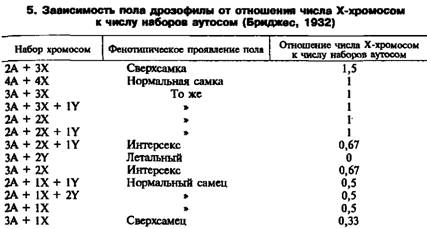
Существует равная вероятность закладки особи мужского или женского пола, так как число сперматозоидов у самцов (яйцеклеток у птиц) с Х- и Y-хромосомами одинаково. Самцов обычно рождается на несколько процентов больше, чем самок, но в ходе эволюции выработался механизм сохранения числового соотношения полов, поэтому к возрасту половой и физиологической зрелости это соотношение выравнивается вследствие более высокой смертности самцов. В процессе индивидуального развития (онтогенеза) происходит дифференциация пола — формирование первичных и вторичных половых признаков, которые приводят к возникновению полового диморфизма, т. е. различию между особями противоположных полов по внешним признакам (пропорции телосложения, масса, окраска шерсти, перьев, наличие или отсутствие вымени, различие в строении половых органов и т. д.). Так, самцы сельскохозяйственных животных всех видов крупнее самок, имеют более мужественный вид — массивную голову, передняя часть туловища у них развита сильнее задней и т. д. У самок лучше развита задняя часть туловища, выражены органы, связанные с осуществлением детородной функции и выкармливанием приплода. У животных нередко нарушено развитие признаков пола. Важный вклад в объяснение причин такого рода патологии внесли исследования Бриджеса и Гольдшмидта. Изучая явления первичного и вторичного нерасхождения половых хромосом у мухи-дрозофилы, Бриджес обнаружил у них изменения первичных и вторичных половых признаков, что зависело от числа половых хромосом и их отношения к числу аутосом в кариотипе (табл 5). Интерсексуальные особи имели переходные формы между самцами и самками; «сверхсамки» и «сверхсамцы» характеризовались резко выраженными женскими или мужскими признаками. На основании результатов этих исследований Бриджес пришел к выводу, что пол особи определяется отношением числа Х-хромосом к числу аутосом (А). У нормальных самок отношение числа X: А равно 1, а у самцов 0,5. Эксперименты Бриджеса показали, что у дрозофил Х-хромосома не содержит генов, определяющих пол. Исходя из этих данных, Бриджес сформулировал теорию пола, суть которой состоит в том, что развитие половых признаков зависит от баланса генов, контролирующих их развитие.
Нарушения в системе половых хромосом. В процессе изучения кариотипов животных были выявлены аномалии в системе половых хромосом, связанные в основном с нарушениями расхождения в мейозе, а также в результате обмена клетками разных индивидуумов. В таблице 6 дана сводка таких нарушений и их связь с фенотипом. Как видно из ее данных, нарушения в системе половых хромосом приводят к резко выраженным нарушениям воспроизводительных функций у животных. Половой хроматин. В ядрах интерфазных клеток у нормальных особей женского пола очень часто обнаруживают небольшую глыбку хроматина, лежащую у ядерной оболочки или представленную в форме барабанной палочки, прикрепленной тонкой нитью к ядру. Барр и Бертрам, впервые обнаружившие это тельце, назвали его «сателлитом ядра». Лайон установила, что эта глыбка хроматина представляет собой одну из двух Х-хромосом, которая находится во время интерфазы в гетеропикнотическом состоянии. Положительный гетеропикноз, по гипотезе Лайон, свидетельствует о генетической инактивации одной из Х-хромосом кариотипа женского пола. Ввиду того что это тельце встречается только в ядрах клеток самок, его рассматривают как признак, отличающий клетки самок от клеток самца, и называют половым хроматином или тельцем Барра.
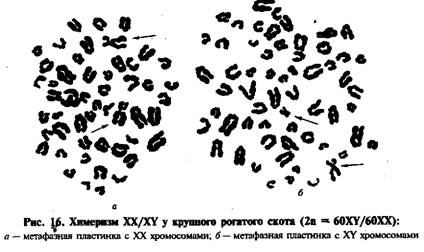
Этот признак можно использовать для изучения нарушений в системе половых хромосом, в диагностике пола при интерсексуальности. Количество телец Барра всегда на единицу меньше числа Х-хромосом. Так, если у самок обнаруживается два тельца Барра, то они являются носителями трисомии по Х-хромосоме. Если половой хроматин отсутствует, то у особи женского пола имеется только одна Х-хромосома. Если у самца обнаруживают тельце Барра, это значит, что у него в кариотипе не одна, а две Х-хромосомы. Для исследования полового хроматина используют клетки лейкоцитов, слизистых полостей и др. Интерсексуальность у животных. Влияние внутренней среды организма на изменение признаков пола в онтогенезе прослеживается при изучении интерсексуальности. У домашних животных существуют разные формы интерсексуальности, которые объединяются под названием гермафродитизм. Кроме того, обнаружены отдельные формы, очень сходные с известными синдромами Клайнфельтера и Тернера. Образование гермафродитов — особей, имеющих гонады и (или) половые органы противоположного пола, рассматриваются как результат нарушения мейоза в период развития бластоцисты. Синдром Клайнфельтера характеризуется недоразвитием гонад, повышением выделения гонадотропина и другими изменениями. Классической формой кариотипа синдрома Клайнфельтера является 2п (аутосом) + XXY. Однако часто в кариотипе обнаруживают мозаицизм по половым хромосомам: XXY/XY/XX и трисомию XXX (синдром «трипло-Х»). У крупного рогатого скота XXY-синдром наблюдали в сочетании с Х-трисомией. Характерными признаками животных были нарушения роста и развития, двусторонняя гипоплазия семенников с олиго- и некроспермией и другими изменениями. Возникновение синдрома Клайнфельтера связано с нерасхождением половых хромосом в мейозе. Примечательной особенностью фенотипа при этом синдроме является то, что животные при наличии даже двух Х-хромосом и одной Y-хромосомы имеют признаки мужского пола, хотя и стерильные. При отсутствии Y-хромосомы животные имеют женский фенотип. Это указывает на то, что Y-хромосома у животных содержит гены, контролирующие развитие мужского пола и плодовитость. Синдром Тернера характеризуется женским фенотипом с дисгенезией гонад и другими генитальными дефектами. Цитогенетически синдром Тернера определяется формулой 2л (аутосом) + ХО (отсутствует вторая Х-хромосома). Животных с признаками гермафродитизма, а также носителей синдромов Клайнфельтера и Тернера следует своевременно выделять в группы откорма. Фримартинизм — особая форма интерсексуальности, выявляемая у крупного рогатого скота. Наблюдения показывают, что бесплодные телки — фримартины рождаются в двойне с бычком. Установлено, что в среднем 95 % телок из разнополых двоен имеют признаки фримартинизма. У них часто обнаруживают мужской тип экстерьера, недоразвитие матки и др. Для объяснения причин бесплодия телок - фримартинов ученые выдвигают разные теории. Основные из них — гормональная и клеточная. Гормональная теория предполагает возникновение фримартинизма вследствие слияния плодного мешка при беременности двойней и образования анастомозов между плацентарными сосудами разнополых плодов, по которым осуществляется обмен мужскими гормонами — андрогенами (тестостерон) и женскими — эстрогенами. Поскольку мужской гормон тестостерон начинает продуцироваться раньше, то длительное воздействие на женские половые органы приводит к вирилизации (недоразвитию) последних, отклонениям в сторону мужского пола и рождению, наконец, бесплодных телок. Клеточная теория исходит из того, что при плацентарных анастомозах происходит обмен не только гормонами, но и другими элементами. Так, у телок - фримартинов был обнаружен химеризм по эритроцитарным антигенам и по половым хромосомам (рис. 16). Химеризм по половым хромосомам наблюдается и у быков из разнополых двоен. У бычков на племпредприятиях частота химеизма в среднем составляла 1,5 %, у отдельных из них — свыше % (А И. Жигачев и др., 1989). Эти животные часто имеют нарушения воспроизводительной функции — от снижения количества спермиев в эякуляте и пониженной оплодотворяющей способности до полного бесплодия.
Степень нарушения плодовитости связана, очевидно, с временем образования сосудистых анастомозов во время внутриутробного развития быков химериков. Химеризм по половым хромосомам наблюдается не только в двойнях, но и в отелах коров с большим числом телят разного пола. Присутствие Y-хромосомы в кариотипе телок-фримартинов и изменение у них признаков в сторону мужского пола — явное свидетельство ее влияния на половые признаки. Цитогенетический анализ — надежный метод ранней диагностики фри-мартинизма у телок. Выявление химеризма XY/XX-хромосом у быков — это критерий малопригодное их в воспроизводстве. Химеризм по половым хромосомам обнаружен также у коз, овец, свиней, норок. В некоторых породах коз (зааненская, тогденбургская) интерсексуальность встречается довольно часто — 6,5—8,4 %. Этот признак связан с животных. По данным Г. И. Исаевой и Д. К. Беляева, химеризм наиболее часто регистрировали у норок, гомозиготных или гетерозиготных по генам алеутской окраски или хедлунд. Причины более высокой смертности среди самцов млекопитающих можно объяснить исходя из особенностей наследования признаков, сцепленных с полом. Явление сцепленного с полом наследования впервые открыл Т. Морган при скрещивании мух-дрозофил с красной и белой окраской глаз. Если в скрещивании участвовали красноглазые самки и белоглазые самцы, все потомство рождалось красноглазым. Во втором же поколении наблюдалось расщепление в соотношении 3:1. Но в отличие от мендельского моногибридного скрещивания расщепление было только среди самцов: одна половина из них была с белыми глазами и другая — с красными; все самки были красноглазыми. При обратном (реципрокном) скрещивании белоглазых самок с красноглазыми самцами картина была иной: все самки имели красные глаза, самцы — белые. При скрещивании этих особей во втором поколении половина самцов и самок рождалась красноглазыми. Признаки, расщепление по которым при скрещивании связано с полом, называют сцепленными с полом. Эти признаки обусловливаются генами, локализованными в половых хромосомах. Установлено, что наследование их зависит в основном от Х-хромосомы Y-Хромосома имеет небольшие размеры, состоит преимущественно из гетерохроматина и является генетически инертной, за исключением, возможно, некоторых генов, контролирующих воспроизводительную функцию и признаки пола. У самцов млекопитающих гены, локализованные в Х-хромосоме, не имеют доминантных или рецессивных партнеров (аллелей) на Y-хромосоме. Рецессивные гены у них проявляют свое действие уже в одинарной дозе (гемизиготном состоянии) по типу доминантного. Практическое использование сцепленного с полом наследования признаков. В птицеводстве оказалась полезной рецессивная, сцепленная с Х-хромосомой мутация карликовости. Карликовые куры отличаются от нормальных лучшей оплатой корма продукцией, для них требуется меньшая площадь содержания. Они резистентны к отдельным болезням. В шелководстве получил распространение метод использования сцепленных с полом деталей для получения гусениц только мужского пола, дающих более крупные коконы, содержащие Шелка на 25—30 % больше, чем коконы гусениц самок. Для этого В. А. Струнников вывел линию, сбалансированную по двум Z-деталям с помощью транслокации (переноса) фрагмента половой Z-хромосомы на W-хромосому При скрещивании самцов с одной неаллельной деталью в каждой из двух хромосом (а и Ь) с самками без транслокаций рождаются только самцы:
ми в половой хромосоме. Как правило, они имеют рецессивный характер проявления, и при этом поражаются преимущественно особи гетерогамного пола: у млекопитающих — самцы, у птиц — самки. Сцепленные с полом летальные и сублетальные аномалии изменяют численное соотношение полов при рождении или после него вследствие гибели или браковки у млекопитающих самцов, у птиц — самок. Например, А. И. Жигачевым установлено, что такая аномалия, как врожденная деформация передних конечностей в сочетании с анкилозом суставов, изученная у животных черно-пестрой, сычевской и костромской пород, проявляется, как правило, у бычков, родственных между собой, что указывает на сцепленное с полом наследование. Наследование признаков, контролируемых генами, локализованными в аутосомах, но фенотипически проявляющихся исключительно или преимущественно у одного пола, — есть наследование, ограниченное полом. Расщепление по таким признакам соответствует правилам Менделя. Выделяют также признаки, контролируемые полом. Они обусловлены генами, локализованными в половых хромосомах, но проявляются у обоих полов по-разному. К числу патологических, или нежелательных, признаков, ограниченных полом, у животных можно отнести гипоплазию вымени и сосков, гипоплазию гонад (семенников и яичников), крипторхизм, недоразвитие разных частей половых органов у телок (наблюдаемое преимущественно у животных белой масти), сужения прямой кишки и матки у коров, аплазию сегментных протоков Вольфа, затянувшуюся беременность у самок, аномалии сперматозоидов у самцов и др. Большинство из указанных аномалий вызывает частичное или полное нарушение воспроизводительной функции . Проблема регуляции пола вытекает из необходимости увеличения продукции животноводства за счет преимущественного получения особей одного вида, дающих более высокий выход молока, мяса, шерсти, яиц и т. д. Так, в молочном скотоводстве более желательно рождение телочек, а в мясном — бычков, так как они быстрее растут. От высокоценных племенных быков и коров целесообразно получать мужских потомков для более быстрого размножения их генотипов. В яичном птицеводстве экономически более выгодно получение курочек. В связи с этими практическими потребностями исследователи не только стремятся познать механизмы определения пола, но и изучают возможности искусственного регулирования пола. Необходимо отметить, что в отношении крупных животных с внутриутробным развитием плодов эта проблема еще не решена. Регуляция соотношения полов у млекопитающих может быть достигнута путем разделения спермы на две фракции: первую — содержащую в спермиях Х-хромосому и вторую — содержащую Y-хромосому. Оплодотворение самок одной из этих фракций будет давать приплод одного пола. Проводились эксперименты по разделению спермы на указанные фракции центрифугированием, электрофорезом и седиментацией (осаждением) с помощью аминокислот гистидина Осеменение самок крольчих, например, более легкой и более подвижной фракцией приводило к сдвигу в сторону мужского пола. Однако полного сдвига в соотношении полов сделано не было. Разрабатывается метод количественного определения ДНК в спермиях путем измерения интенсивности флуоресценции ядер. Полученные результаты, как считают авторы этого метода, могут стать предпосылкой для успешного разделения спермиев у млекопитающих на несущие X-или Y-хромосому. Партеногенез. Это развитие организма без оплодотворения. Получение особей одного пола может быть достигнуто при развитии эмбрионов из отцовских (андрогенез) или материнских (гиногенез) гамет. Так, под руководством Б. Л. Астаурова были проведены эксперименты по андрогенезу у тутового шелкопряда. Неоплодотворенные яйца шелкопряда подвергали тепловому шоку и облучали рентгеном, тем самым разрушали их ядра, не повредив цитоплазму. Затем эти яйца осеменяли. Зигота формировалась путем слияния ядер двух проникших в яйцо спермиев, развившиеся из нее особи имели признаки только отцовского вида. В другом варианте экспериментов неоплодотворенные и еще не прошедшие редукционного деления яйца нагревали, останавливая тем самым мейоз и сохраняя диплоидность набора хромосом. Из таких яиц без оплодотворения (партеногенетически) развивались только самки, унаследовавшие признаки матери. Девственное развитие (партеногенез) в естественных или спонтанных условиях встречается у птиц. И. В. Кудрявцев, 3. А. Ощепкова, А. К. Голубев и др. на основании экспериментов, проведенных на курах разных пород, пришли к выводу, что существуют генетические предпосылки селекции на получение жизнеспособных особей — партеногенов и создание линий с высокой предрасположенностью к партеногенезу. При этом все вылупляющиеся цыплята оказываются петушками. Отбор на повышение частоты партеногенеза, проведенный в двух линиях индеек, дал следующие результаты. Способность яиц к партеногенезу возросла с 1,1 до 18,6 % в первой линии ис4до21,1%во второй. Значение партеногенеза не только в том, что он позволяет получить потомство одного пола, но и в том, что использование этого явления дает возможность получить особей, идентичных генотипу одной из родительских форм.
Билет 18
Наследование, сцепленное с полом — наследование какого-либо гена, находящегося в половых хромосомах. Наследование признаков, проявляющихся только у особей одного пола, но не определяемых генами, находящимися в половых хромосомах, называется наследованием, ограниченным полом. Наследованием, сцепленным с X-хромосомой, называют наследование генов в случае, когда мужской пол гетерогаметен и характеризуется наличием Y-хромосомы (XY), а особи женского пола гомогаметны и имеют две X-хромосомы (XX). Таким типом наследования обладают все млекопитающие (в том числе человек), большинство насекомых и пресмыкающихся. Наследованием, сцепленным с Z-хромосомой, называют наследование генов в случае, когда женский пол гетерогаметен и характеризуется наличием W-хромосомы (ZW), а особи мужского пола гомогаметны и имеют две Z-хромосомы (ZZ). Таким типом наследования обладают все представители класса птиц. Если аллель сцепленного с полом гена, находящегося в X-хромосоме или Z-хромосоме, является рецессивным, то признак, определяемый этим геном, проявляется у всех особей гетерогаметного пола, которые получили этот аллель вместе с половой хромосомой, и у гомозиготных по этому аллелю особей гомогаметного пола. Это объясняется тем, что вторая половая хромосома (Y или W) у гетерогаметного пола не несет аллелей большинства или всех генов, находящихся в парной хромосоме. Таким признаком гораздо чаще будут обладать особи гетерогаметного пола. Поэтому заболеваниями, которые вызываются рецессивными аллелями сцепленных с полом генов, гораздо чаще болеют мужчины, а женщины часто являются носителями таких аллелей. Примеры заболеваний человека, сцепленных с полом[править | править вики-текст] · Гемофилия A · Гемофилия В · Дальтонизм · Лекарственная гемолитическая анемия, связанная с дефицитом глюкозо-6-фосфатдегидрогеназы (Г6ФД) · Синдром Леша-Найхана · X-связанный ихтиоз
Билет 19
Биологическая роль нуклеиновых кислот заключается в том, что ДНК хранит наследственную информацию организма в виде последовательности дезоксирибонуклеотидов, различающихся азотистыми основаниями. В ДНК в закодированном виде записан соста всех белков организма. Каждой аминокислоте, входящей в состав белков, соответствует свой код в ДНК, а именно - три конкретных нуклеотида. Молекулы РНК переносят информацию от ДНК к местам клетки, где происходит синтез белка. Первичная структура НК - это последовательность расположения мононуклеотидов в полинуклеотидной цепи (рис. 4.4.1.).
Вторичная структура - молекула, состоящая из двух (ДНК) или одной (РНК) правозакрученных вокруг воображаемой оси спиралей. В ДНК направление фосфотиэфидных связей(3'-5') антипараллельно. Третичная структура НК - это кольцо из ДНК, что имеет место у бактерий и вирусов. Функции нуклеиновых кислот рассмотрены в главе "Матричный биосинтез". Дата добавления: 2015-12-16 | Просмотры: 4462 | Нарушение авторских прав |