|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
КЛЮЧЕВЫЕ ПОЗИЦИИ
Ключевыми положениями тибетской медицинской системы являются два момента: 1) три системы регуляции состояния организма и 2) понятия о тепле и холоде, горячих и холодных болезнях. Целью нашей работы является идентификация трех тибетских систем регуляции состояния организма с современных позиций на основании сопоставления тибетской и современной информации по механизмам регуляции. Мы попытались также интерпретировать представления тибетской медицины о тепле и холоде, горячих и холодных болезнях. По представлениям тибетской медицины, состояние организма человека регулируют три системы: первая называется «рлунг» (дословный перевод — ветер), вторая — «мкхрио (желчь) и третья — «бадкан» (слизь). Исходя из названия систем, можно предполагать, что каждая из систем имеет специфические способы и факторы действия. Так, фактор регуляции, используемый в первой системе, подобен ветру, он распространяется, перемещается, как ветер. Вторая система регуляции — желчь, очевидно, пользуется способом регуляции, который подобен способу распространения желчи. Используемый этой системой фактор регуляции также подобен желчи, т. е. вырабатывается в клетках тканей, как желчь в клетках печени, распространяется по сосудам, как желчь, и попадает в различные органы и ткани, где и действует как фактор регуляции. Третья система — слизь, возможно, имеет фактор регуляции, подобный слизи, который.образуется в тканях и реализует свое влияние в месте образования, т. е. эта система является местнотканевым способом регуляции. Подобные интерпретации каждой из трех систем регуляции по факторам и способам регуляции, очевидно, адекватны, однако они являются лишь частичными характеристиками систем регуляции. Направленность и результаты функционирования каждой из трех систем являются следующими важными их параметрами. Внутри каждой из трех тибетских систем регуляции различают по пять разновидностей, подсистем, которые имеют свои специфические функции в организме. Каждая из пяти разновидностей (подсистем) системы ветра, а также и двух других систем, желчи и слизи, имеет названия, соответствующие тем функциям, которые они выполняют в организме. Разновидности системы ветра имеют следующие названия и участвуют в регуляции перечисленных ниже процессов: 1) жизненный, или поддерживающий жизнь, регулирует функционирование сердечно-сосудистой системы, дыхание; 2) движущийся вверх — участвует в развитии всех процессов, происходящих в верхней части тела, в области головы; 3) проникающий всюду — регулирует разнообразные процессы в тканях и органах всего тела; ровный огонь — участвует в регуляции пищеварения и усвоении пищи; очищающий вниз — определяет выделение нечистот из организма (кала и мочи), функционирование половой сферы.
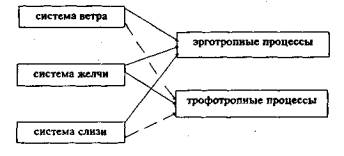
1) пищеварительная — участвует в пищеварении; 2) меняющая цвет — участвует в усвоении поступающего в организм, в кроветворении, определяет окраску кожных покровов и других тканей; 3) сопутствующая — регулирует функции сердечно-сосудистой системы, определяет ясность мысли, гордость, решительность, различные желания; 4} дающая зрение — дает возможность видеть; 5) чистящая цвет кожи — определяет чистоту кожных покровов. Пять разновидностей системы слизи и их функции в организме приведены ниже: 1) поддерживающая — дает жизнь; 2) смешанная — участвует в пищеварении; 3) испытующая — определяет формирование вкусовых ощущений; 4) удовлетворяющая — регулирует функционирование всех органов чувств, участвует во всех процессах, происходящих в мозге; 5) соединяющая — «соединяет конечности», определяет чувствительность И движения в них. Каждая из трех систем регуляции имеет свои центральные и периферические структурные образования — органы, а две системы — ветер и желчь, также имеют сосудистую сеть. Местоположение центральных структурно-морфологических образований, органов и их функций по каждой из трех систем регуляции просматривается, начиная с эмбрионального периода, со времени его закладки. Так, центральный орган системы ветра у эмбриона закладывается в пояснично-крестцовой области, в тазу. У взрослого человека центральный орган этой системы регуляции также находится в поясничной области. Центральный орган второй регулирующей системы организма — желчи — находится в печени эмбриона, а затем, после рождения, система желчи, имея центральное местоположение в печени, распространяется по организму и присутствует практически во всех тканях взрослого человека. Центральный орган третьей системы регуляции слизи закладывается у эмбриона в мозгу и затем на протяжении жизни это структурное образование в мозгу продолжает оставаться центральным органом этой системы! Система слизи также имеет представительство во всех тканях организма. Каждый из пяти подвидов системы регуляции имеет основное местоположение, однако конкретные структурные образования в этих местоположениях в тексте «Чжуд-ши» не указываются. Ветер и желчь, факторы регуляции этих систем, распространяются в организме повсеместно, проникают всюду, циркулируют по сосудам четырех видов, белым, желтым, красным и черным. Судя по цвету сосудов, а также по схемам, представленным на картинах «Атласа тибетской медицины», сосуды белые являются нервными стволами, нервами, сосуды желтые — лимфатическими, а красные и черные.— кровеносными сосудами. Третья система регуляции, как уже было отмечено, использует фактор регуляции, который действует местно в тех тканях, где он образуется, поэтому система слизи не имеет своей специфической сосудистой сети. Европейская медицина, как известно, различает только две системы регуляции состояния организма, нервную и гуморально-эндокринную. Различные аспекты строения и функционирования нервной и гуморально-эндокринной систем регуляции достаточно хорошо исследованы, этим вопросам посвящена обширная литература. В настоящей главе приводится лишь та, небольшая часть литературной информации, посвященной системам регуляции, их механизмам, веществам и эффектам, которая была необходима для идентификации тибетских систем регуляции. В последние два десятилетия сведения о регуляторных системах организма, нервной и гуморально-эндокринной, значительно пополнились и вместе с этим представления о них существенно изменились. Изменения в представления о механизмах нервной и гуморально-эндокринной регуляции внесли данные о роли пептидов в регуляторных процессах. Изучение пептидов проводится в течение почти столетия, однако лишь информация, полученная в 70—80-е годы, позволила, хотя и далеко еще не полностью, установить их важную роль в механизмах регуляции различных процессов, происходящих в организме. Пептиды представляют особый класс активных регуляторных соединений, участвующих в определении состояния различных органов и тканей организма. Полипептиды, по определению Фишера (Fischer, 1906),— соединения, образуемые аминокислотами и имеющие в своем составе амидные связи, регулярно повторяющиеся связи аминокислотных остатков. По числу аминокислот в пептидной цепи различают ди-, три-, тетра-пептиды и т. д. Пептидные цепи могут спонтанно укладываться в сложные трехмерные структуры, что является характерной особенностью белков. В настоящее время число известных регуляторных пептидов составляет несколько десятков и численность их продолжает увеличиваться, среди них: окситоцин, вазопрессин или антидиуретический гормон, нейрофизин, кортикотропин или адренокортикотропин (АКТГ), метионинэнкефалин, лейцинэнкефалин, а-меланотропин, соматотропин или гормон роста (СТГ), соматолиберин, соматостатии, кортиколиберин, тиролиберин, люлиберин, пролактолиберин, пролактостатин, меланолиберин, меланостатин, субстанция Р, эледоизин, физалемин, бомбезин, нейротензин, ангиотензин I, ангиотензин II, ангиотензин III, каллидин, брадикинин, пептиддельта сна, панкреатический полипептид, холецистокинин-панкреозимин, гастрин, гастринрилизинг пептид, глюкагон, инсулин (А цепь), инсулин-рилизинг пептид, вазоактивный кишечный пептид (ВКП, ВИП). Предлатается также включить в этот перечень регуляторных пептидов у-аминобутирил, уротензин, проктолин, тимопоэтин, тафцин, лейкоцитарный интерферон и др. Перечисленные пептиды обнаружены в ткани мозга и в тканях внутренних органов. В ткани мозга содержатся следующие пептиды: окситоцин, вазопрессин, липотрофин, АКТГ, Д-эндорфин, а-эндорфин, у-эндорфин, метэнкефалин, лей5-энкефалин, а-меланотропин, меланотропин, у-меланотропин, СТГ, соматолиберин, соматостатин, кортиколиберин, соваган, тиролиберин, люлиберин, гонадолиберин, пролактолиберин, пролактостатин, меланолиберин, меланостатин, субстанция Р, эледоизин, физалемин, бомбезин, ранатензин С и ранатензин В, алитензин, церулоплазмин, церулеин, нейротензин, ангиотензин II и ангиотензин III, каллидин, брадикинин, панкреатический полипептид, холецистокинин-панкреозимин, гастрин, гастрин-рилизинг пептид, секретин, глюкагон, инсулин-рилизинг пептид, инсулин, ВИП, гамма-аминомасляная кислота (ГАМК). Перечень пептидов, обнаруживаемых в клетках мозга, продолжает увеличиваться. Как следует из приведенных данных, в число пептидов мозга входят классические гормоны, участвующие в функционировании системы пищеварения: гастрин, инсулин, холецистокинин-панкреозимин, глюкагон и др. Как известно, существуют нейроны, секретирующие адреналин, норадреналин (адренэргические) и продуцирующие ацетилхолин (холинэргические). По мере детального исследования нейрональных секретов было установлено существование пуринэргических (Burnstock, 1972), а также пептидэргических нейронов (Larsson, 1980). Вместе с тем, многие исследователи выражают сомнение в существовании пуринэргических нейронов (Bloom, Polak, 1978, Humphrey, Fischer, 1978, Jones, 1978, и др.) и приводят доводы в пользу существования только трех типов нейронов: адренэргических, холинэргических и пептидэргических (Larsson, 1980). Кроме того, показано, что «в одном нейроне одновременно с классическими нейромедиаторами могут находиться регуляторные пептиды, например, в клетках миндалевидного комплекса, в нейронах шва продолговатого мозга, а также в нервных терминалях в мозге обнаружены норадреналин и энкефалин, норадреналин и соматостатин, или серотонин и субстанция Р и т. д. (Hokfelt at al., 1978a, 19786). Вследствие выяснения подобных ситуаций несостоятельной оказалась принятая ранее концепция Дэйла (1953) (цитировано по П.К.Климову, 1986) «один нейрон — один трансмиттер». Деятельность мозга основана на объединении нервных клеток в целостную систему, в которой существуют различные виды связи, взаимодействия клеток внутри самого мозга и с различными висцеральными системами и тканями организма. В пределах мозга связи различных нейронов, разных областей и структурных образований могут осуществляться нейромедиаторным путем, а также гуморально-эндокринным дистантным и паракринным, местно-тканевым, способами. Представления о нейромедиаторной регуляции в целом и в частности на уровне самого мозга, его структур, в значительной мере изменились. «Простая и удобная схема для построения теории регулирования, по которой в центральной нервной системе действуют несколько нейромедиаторов, одни из которых передают возбуждение, другие его тормозят, оказалась сломленной под натиском новых фактов». (П.К.Климов, 1986). К числу подобных новых фактов относятся данные о том, что адренэргические и холинэргические межнейронные связи в ткани мозга составляют не более 15% всех синапсов (Nicoll, 1976, Snyder at al., 1976). Установлено, что нейротрансмиттерные функции, кроме классических нейромедиаторов — ацетилхолина и норадреналина, могут выполнять и другие соединения, такие как аденозинтрифосфатная кислота (АТФ), серотонин, гистамин, дофамин (в симпатических узлах млекопитающих), пептид субстанция Р (в сенсорных нервных волокнах) (Ноздрачев, 1983). Предполагается, что и некоторые другие пептиды могут выступать в качестве нейротрансмиттеров в межнейрональных связях. И. П. Ашмарин (1982) полагает, что значительная часть известных нейропептидов (бомбезин, холецистокинин, вазоактивный кишечный пептид, нейротензин, энкефалины, субстанция Р, тимозин и др.) могут быть нейротрансмиттерами, так как отвечают всем требованиям, предъявляемым к таковым. Так, энкефалины обнаружены в аксонах и нервных окончаниях со многими разветвленностями, имеющими соприкосновение с сетью терминалей нейронов, содержащих адреналин, дофамин, серотонин (Snyder, 1979). Места локализации энкефалиновых терминалей нейронов совпадают с местонахождением опиатных рецепторов. Волокна, содержащие энкефалин, есть в таламусе, бледном шаре, полосатом теле, также содержат энкефалиновые нервы хвостатое ядро, миндалевидный комплекс, особенно его центральное ядро, меньше таких волокон в коре и гипофизе. Нейропептиды, выполняющие функции нейротрансмиттеров, по нейрональным отросткам доходят до клеток-мишеней в мозге, выделяются в синаптическую щель, где и связываются с постсинаптической мембраной клетки-мишени. Пептиды в этом случае могут выполнять роль нейротрансмиттеров для относительно медленной передачи сигналов в центральной нервной системе. Не исключается также возможность того, что в синаптической щели они могут действовать на пре- или постсинаптические рецепторы с тем, чтобы модулировать состояние активности или торможения для облегчения или подавления синаптической передачи в интернейрональной связи (Stewart, Channabasavaian, 1979). Установлено участие в интернейрональной связи вазоактивного кишечного пептида, нейротензина, субстанции Р, октапептида холецистокинина (CCK8), гастрина, тиролиберина, люлиберина и др. В настоящее время еще мало получено информации о нейротрансмиттерном использовании регуляторных пептидов на уровне мозга, в центральной нервной системе. Значительно больше литературных данных о функционировании их в качестве нейротрансмиттеров в автономной (периферической) нервной системе. Тем не менее, предположения о нейромедиаторной функции некоторых пептидов в мозге имеют фактическое обоснование. Пептиды, секретируемые клетками мозга, могут также функционировать как гормоны в пределах мозга при выделении их в кровеносное русло. Так, например, известно, что пептиды гипофиза тиролиберин и люлиберин, которые могут выступать и в роли нейротрансмиттеров (при передаче регуляторного сигнала клеткам срединного возвышения гипоталамуса), преимущественно функционируют как гормоны, достигая с кровотоком эндокринных клеток гипофиза. В мозге наиболее богаты пептидами нейроны гипоталамуса, миндалевидного комплекса и подкорковых ганглиев. Между этими структурами мозга существуют и нейрональные и эндокринные связи, которые крайне важны как для функционирования самого мозга, так и в определении состояния висцеральных органов (Баклаваджян, 1969, Черкес, 1969, Ажипа, 1979, Баранова, 1979, Константинова, 1979, С.С. Мусящикова и В.Н.Черниговский, 1973). На уровне мозга межнейронные контакты могут также осуществляться паракринным путем, через открытые межклеточные синапсы. Так, диффузия пептидов по межклеточным пространствам обнаружена в таких структурах мозга как базальные ганглии, новая кора, III желудочек (Krisch, Leenhardt, 1980). Передача информации от нейрона к нейрону при этом также может происходить с помощью широко распространенных в ткани мозга разнообразных пептидов. При паракринном взаимодействии различные виды секреторных клеток могут выделять в околоклеточную щель свои секреты — биологически активные вещества, т. е. создается при этих условиях своего рода открытый широкий синапс, который обеспечивает одновременное присутствие нескольких видов регуляторных сигналов. Естественно, что подобный открытый синапс не является жестко фиксированным по составляющим структурным компонентам, и как следствие этого выступает разнообразие физиологических процессов, развивающихся в различных тканях и даже в.пределах одной и той же ткани. В то же время межнейронные синапсы имеют жесткие анатомические пути, передающие и принимающие звенья регуляторной информации, передаваемой с помощью определенных нейротрансмиттеров, и как следствие этого — развиваемые ими более или менее однозначные физиологические эффекты. Механизмы действия гуморально-эндокринных регуляторов, транспортируемых через кровоток, и соответственно вызываемые ими физиологические эффекты, очевидно, занимают промежуточное положение между весьма разнообразными по виду и варьирующими по месту и времени развития физиологическими эффектами, развивающимися вследствие регуляции через местно-тканевые открытые межклеточные синапсы, и достаточно жестко закрепленными по времени и месту реализации, по путям передачи, по характеру физиологического эффекта нейромедиаторными регуляторными сигналами. Биологически активные соединения, выделившиеся в околоклеточную синаптическую щель, могут вступать в непосредственную связь с рецепторами клеток, ограничивающих околоклеточное пространство, или могут вступать во взаимодействия с другими биологически активными соединениями, находящимися в околоклеточном пространстве, следствием чего может быть видоизменение регуляторного сигнала, его ослабление, усиление или нейтрализация. Общеизвестно широкое распространение в тканях системы пищеварения клеток, содержащих регуляторные пептиды. Содержание таких пептид-секретирующих или пептид-содержащих клеток в других тканях исследовано в меньшей мере. В слизистой оболочке кишечника в основе крипт и ворсинок обнаружены клетки, которые названы энтеро-хромафинными (ЕС), содержащие серотонин и пептиды (субстанцию Р, энкефалины, мотилин). Среди ЕС клеток выделяют ЕС1-клетки, содержащие серотонин и субстанцию Р, находятся они в кишечнике и поджелудочной железе, ЕСа-клетки, содержащие серотонин и мотилин, находятся в слизистой толстого кишечника и в поджелудочной железе, ЕС2-клетки, содержащие серотонин, обнаружены они в эпителии слизистой желудка и тонкого кишечника (Grobe U., Forssman W.G., 1979). В слизистой же желудка, тонкого кишечника, поджелудочной железы находятся Д-клетки, содержащие соматостатин, мет-5-энкефалин, гастрин, в других клетках, которые называют Дг или Н-клетки, обнаружен ВИП, эти клетки встречаются в кишечнике, желудке и поджелудочной железе. А-клетки поджелудочной железы, кишечника и желудка содержат целый ряд пептидов: глюкагон, глисентин, холецистокинин-панкреозимин, а также, возможно, эндорфины. S-клетки кишечника содержат секретин, дофамин и холецистокинин. В К-клетках тонкого кишечника присутствует желудочный тормозной пептид. Только нейротензин обнаружен в N (1)-клетках слизистой желудка, двенадцатиперстной кишки и кишечника. Р-клетки желудка, тонкого кишечника и, возможно, поджелудочной железы секретируют бомбезин, а РР(Ф)-клетки тонкого кишечника, желудка и поджелудочной железы хранят панкреатический полипептид и мет5-энкефалин. Из приведенной информации следует, что, по меньшей мере, 17 пептидов находятся (секретируются или хранятся) в клетках тканей системы пищеварения. Пептиды, содержащиеся в клетках тканей пищеварительной системы, возможно, реализуют регуляторное действие в пределах той ткани, в клетках которой они находятся, всеми тремя способами, нейромедиаторным, эндокринным и паракринным (местно-тканевым). Местные тканевые физиологические эффекты перечисленных выше регуляторных соединений описаны в соответствующей литературе (В.Т. Ивашкин, 1981, П.К.Климов, 1983, 1986). Дистантное регуляторное влияние указанные соединения, очевидно, реализуют нейромедиаторным и эндокринно-гуморалъным способами. Эти пути распространения регуляторных соединений, очевидно, являются способами связи с другими висцеральными системами и с мозговыми структурами. При гуморально-эндокринном распространении на пути от места синтеза и высвобождения до мозговых структур регуляторные пептиды должны пройти гематоэнцефалический барьер, на возможность его преодоления пептидами указывают различные исследователи (Прибрам, 1975). Возможно также, пептиды участвуют в роли нейромедиаторов в афферентных сигналах от висцеральных систем, в том числе от пищеварительной, в центральную нервную систему. Единичные наблюдения подобного рода в литературе приводятся. Пептиды обнаружены в чувствительных нейронах слизистой желудочно-кишечного тракта (Бюрнсток, 1981). Известно также участие пептидов (субстанции Р) в передаче афферентных сигналов в качестве нейротрансмиттеров в нервных волокнах, связанных с болевой чувствительностью (Otsuka, Konishi, 1983). В клетках мозга, в различных его структурах, также синтезируются, хранятся, выделяются и действуют те же пептиды или их аналоги, они участвуют в таких процессах как обучение, запоминание, терморегуляция, лактация, регуляция состояний различных функциональных систем организма, пищеварения, дыхания, сердечно-сосудистой и др. Поступление пептидов от тканей различных внутренних органов по нервным путям или по кровеносным сосудам может менять течение внутримозговых процессов, состояние отдельных нейронов или определенных центров, ядер мозга, определяя их активность или торможение, принятие решения и формирование эфферентного сигнала. Как считают некоторые исследователи (П.К.Климов, 1986), «одной из физиологических функций, может быть самой главной, природных пептидов мозга и органов висцеральной системы является участие их в переносе информации о функциональном состоянии органов, а также всей пищеварительной системы. В сущности речь идет о создании мозгом «образа» желудочно-кишечного тракта, «слепка» его функционального состояния, в принятии решения и выработке эфферентных, сигналов». В принятии решения на уровне мозга может участвовать информация, поступившая от других висцеральных органов, от органов чувств, информация зрительного, обонятельного, вкусового характера. Принятое решение по эфферентным нервным путям направляется к исходному органу, к другим органам и ко всей функциональной системе, в данном случае к органам и тканям системы пищеварения. Эфферентные сигналы могут поступать к исходному органу или всей функциональной системе по эфферентным нервным путям и через гуморальную циркуляцию. Эфферентные пути от коры головного мозга к желудку и двенадцатиперстной кишке в настоящее время известны только в общих чертах, они достаточно сложны и включают целый ряд образований, таких как ретикулярная формация, миндалевидный комплекс, гипоталамус (Богач, 1977, 1982). Нервные эфферентные пути, по которым передаются сигналы к желудку и кишечнику, включают блуждающие и чревные нервы. В литературе приводятся многочисленные данные об участии пептидов, наряду с классическими нейромедиаторами, в качестве нейротрансмиттеров в проведении эфферентных регуляторных сигналов к различным тканям системы пищеварения. Участниками синаптической передачи информации являются АТФ, серотонин, ГАМК, дофамин, энкефалины, вазоактивный кишечный пептид, бомбезин, соматостатин, нейротензин, холецистокинин, гастрин, панкреатический полипептид, брадикинин, мотилин (Burastock, 1981), ангиотензин II, тиролиберин (П.К.Климов, 1986). Пептиды обнаружены в интернейронах, в нервах, снабжающих кровеносные сосуды и эпителиальные клетки слизистой оболочки желудочно-кишечного тракта (Burastock, 1981). Среди волокон блуждающего нерва — множество пептидсодержащих волокон (Lundberg et al., 1978; Gamse et al., 1979). Известные в настоящее время механизмы и факторы эндокринной регуляции изложены в соответствующих руководствах {М.И. Балаболкин, 1989, и др.). Клетки, содержащие и выделяющие регуляторные пептиды в головном мозге и других тканях, предложено рассматривать как своеобразную систему регуляции и называть ее «диффузная нейроэндокринная система» (DNES — diffuse neuro-endocrine system) (Pearse, Takor, 1979), «АРИД-система» — в связи со свойствами ее клеток (amine content, precursor uptake, decarboxylation — амин-содержащие, поглощающие предшественников биогенных аминов с последующим их декарбоксилированием; Pearse, Takor, 1979), «регуляторные клетки» (Grossman, 1979). Исследования, посвященные морфологическим особенностям пептидэргических нейронов мозга, нейросекреторных, и обычных «нормальных» нейронов, не содержащих пептиды, показали, что их морфологические и субмикроскопические признаки одинаковы, разница состоит лишь в том, что клеточные органеллы аппарата белкового синтеза у пептидэргических нейронов выражены сильнее и масса структур-переносчиков секретируемых пептидов намного больше, чем у обычных нейронов (Gersch, Richter, 1981). Исследования, посвященные выявлению различий пептидэргических и обычных нейронов по биологической активности, также показали, что весьма трудно разграничить их и по этому параметру, так как пептидэргические нейроны оказывают модулирующее влияние на деятельность нервных клеток головного мозга (Samson, Kozlowski, 1981). Ряд авторов сопоставляли морфологические и функциональные признаки обычных нейронов мозга, сенсорных нейронов вегетативной нервной системы, пептидэргических нейронов мозга и энтеропептидсодержащих клеток (клеток в тканях системы пищеварения). В результате подобного сравнительного изучения исследователи пришли к заключению о том, что все эти клетки чрезвычайно близки, «сходны» (Fujita, Kobayashi, 1973; Fujita, 1976, 1977). В связи с этим, предлагается пептидсодержащие и секретирующие их клетки называть паранейронами, в частности, эндокринные клетки кишечника и поджелудочной железы (Fujita, Kobayashi, 1981). Таким образом, необходимо отметить появление тенденции рассматривать диффузно рассеянные в ткани мозга и висцеральных органах пептидсекретирующие клетки как функционально единую систему, основной функцией которой является, продукция регуляторных пептидов. Возможно, что диффузно рассеянные в тканях мозга и висцеральных систем клетки АРЙД-системы, или диффузной нейроэндокринной системы, составляют морфологическую основу третьей, выделяемой тибетской медициной местно-тканевой системы регуляции, системы слизи. Как было отмечено в начале настоящей главы, каждая из трех систем регуляции, по тибетским представлениям, имеет свои центральные структурные образования, центральные органы. Первая система регуляции — ветер — имеет центральный орган, располагающийся в области поясницы, возможно, это надпочечники, центральным органом второй системы регуляции — желчи — является печень, и центральное органное представительство третьей системы регуляции — слизи — находится в мозге. Как следует из приведенных данных, в различных системах регуляции, дистантных, нервной и гумррально-эндокринной, и местно-тканевой, паракринной, в качестве передатчиков регуляторной информации в ряде случаев используются одни и те же соединения или их аналоги, ацетилхолин, моноамины, АТФ, серотонин, гистамин, разнообразные пептиды. Не исключена возможность того, что по мере дальнейшего изучения механизмов и факторов регуляторных процессов в организме перечень соединений, используемых параллельно в разных системах регуляции, значительно возрастет. В подобных случаях, кроме химической структуры регуляторного вещества — основного фактора, определяющего характер сигнала, особенности регуляторной информации, возможно, обеспечивает способ ее передачи. Так, известно, что нейромедиаторный способ передачи информации определяет точность, а также высокую скорость передачи регуляторного сигнала. Гуморально-эндокринный и местно-тканевой способы передачи информации, возможно, менее целенаправленны и более продолжительны по времени поступления и действия. Не исключена и возможность того, что гуморально-эндокринный и местно-тканевой способы регуляции, особенно последний, в ряде случаев могут быть многозвеньевыми, протекающими с участием нескольких различных биологически активных соединений, что в конечном итоге обеспечивает определенное отличие их от однонаправленных нейромедиаторных сигналов. Разнообразие ответных реакций на поступление регуляторных сигналов обеспечивается разнообразием видов регулируемых клеток и тканей-мишеней, а также различными состояниями клеток-мишеней, выражающимися в особенностях рецепторного восприятия регуляторного сигнала. Физиологическим эффектам, развивающимся вследствие действия указанных регуляторных соединений, посвящены многочисленные публикации. Обсуждение этих вопросов не являлось целью настоящей работы, и, кроме того, этой проблеме должна быть посвящена не одна монография. В связи с этим, мы рекомендуем интересующимся этими вопросами, обратиться к специальной литературе, в которой эти вопросы детально освещены (П.К. Климов, 1986; М.И. Балаболкин, 1989, и др.). Все разнообразные физиологические процессы, развертывающиеся в клетках тканей организма в результате действия регуляторных стимулов, можно сгруппировать в два класса: 1) эрготропные (от греческого ergon — работа, деятельность) — многочисленные и разнообразные, специфические для каждого вида клеток, процессы функциональной активности (продукция специфических секретов, двигательная активность и др.) и 2) трофотропные процессы (от греческого trophe — питание), такие, как питание, накопление энергетических и пластических материалов в клетке, рост и клеточное деление. Две группы процессов, развивающихся в клетках, находятся в определенных взаимоотношениях, они сменяют друг друга. Так, выполнение специфических для клетки эрготропных процессов происходит с затратой, расходованием энергетических и пластических соединений, т. е. в процессе функционирования клетки тканей «изнашиваются», «истощаются», поэтому состояния функционирования клеток должны сопровождаться восстановительными процессами, трофотропными, которые компенсируют клеточные и тканевые затраты, потери, происходящие при эрготропных процессах. Тибетские представления о тепле и холоде мы комментируем следующим образом: состояние тепла — способность клетки, ткани, органа функционировать, состояние функциональной активности, т. е. эрготропные процессы, состояние холода — снижение способности функционировать, утрата этой способности, а также развитие процессов питания, роста, клеточного воспроизведения, т. е. трофотропные процессы. Снижение способности функционировать может быть выражено в разной степени, т. е. холод может быть незначительный, выраженный и сильно выраженный. Характер действия регуляторных соединений, используемых в каждом из способов регуляции (систем регуляции), возможно, находит отражение в терминах «горячий» и «холодный», т. е., по нашим представлениям, стимулирующий эрготропные процессы, и подавляющий их и усиливающий трофотропные процессы соответственно. В первой системе регуляции, ветре, могут действовать «горячий» ветер и «холодный» ветер, следствием их регуляторного действия являются горячий и холодный процессы в регулируемых тканях. Во второй системе регуляции, желчи, также могут использоваться «горячие» и «холодные» регуляторные соединения, и следствием их действия на клетки и ткани-мишени, соответственно, будет развитие горячих и холодных процессов. И в третьей системе регуляции, слизи, регуляторные факторы также могут использоваться и «горячие» и «холодные», и в результате их регуляторного действия также будут развиваться горячие и холодные процессы. Согласно представлениям тибетской медицины, каждая из систем регуляции участвует в развитии горячих и холодных состояний (эрготропных и трофотропных процессов соответственно) в разной мере. Система ветра в большей степени обеспечивает развитие эрготропных (горячих) процессов, система желчи в равной мере обеспечивает развитие горячих и холодных процессов, а система слизи в большей мере причастна к развитию холодных процессов (рис. 1). системы регуляции регулируемые процессы в тканях
Обозначения: ------- активное участие в развитии процессов - - - - незначительная степень участия в развитии процессов Рис. 1 Участие систем регуляции в развитии эрготропных и трофотропных процессов в тканях Возможно, нервная регуляция различных реакций клеток преследует, в основном, интересы целостного организма, а местнотканевой способ регуляции в большей степени выражает интересы местно-тканевые, интересы самой клетки, среди которых первоочередными являются процессы питания, роста, клеточного размножения, а затем уже следуют процессы специализированного функционирования в интересах организма, других его тканей. Гуморально-эндокринный способ регуляции, возможно, в равной мере обеспечивает развитие процессов, отвечающих интересам местно-тканевым и организменным. Нарушения в состоянии систем регуляции приводят к изменениям регулируемых процессов, как эрготропных, так и трофотропных, к развитию заболеваний (см. главы «Классификация болезней», «Причины болезней»). Болезни различают горячие и холодные. Основу горячих болезней составляют чрезвычайно выраженные, сверхвыраженные, горячие процессы, т. е. состояния гиперфункционирования в органе, в ткани, функциональной системе. К группе горячих болезней мы также относим воспалительные процессы, как выражения гиперфункционирования систем защиты организма. В основе холодных болезней лежат гипофункциональные состояния различной степени, а также чрезмерно выраженные процессы питания, роста, клеточного деления. Три системы регуляции участвуют в развитии горячих и холодных патологических состояний в разной мере. Система желчи, ее расстройства — причина появления горячих болезней, нарушения в системе слизи приводят к развитию холодных болезней, система ветра участвует в развитии и горячих и холодных болезней. Таким образом, решающим моментом для определения системы регуляции в тибетской медицине является способ регуляции, передачи регуляторного сигнала. По этому критерию выделены 3 системы регуляции: ветер, желчь и слизь, которые мы идентифицировали как нервный, гуморальный и местно-тканевой способы регуляции соответственно. В каждой из трех систем регуляции функционируют переносчики регуляторной информации двух видов: горячие и холодные. В тканях организма также происходят горячие и холодные процессы. Горячие процессы идентифицированы нами как состояния функционирования, функциональной активности, а процессы холодные как противоположные состояния, т.е. снижение способности функционировать, функциональной активности и развитие процессов питания, роста и клеточного деления. Горячие регуляторные факторы используются во всех трех системах и действие их вызывает развитие горячих процессов в тканях. Холодные регуляторные факторы также используются всеми тремя системами регуляции и вызывают они развитие в тканях холодных процессов. Действие трех систем регуляции определяет развитие не только физиологических горячих и холодных процессов, но и возникновение горячих и холодных болезней. При обсуждении последующих вопросов мы будем использовать тибетские названия трех систем регуляции: ветер, желчь, слизь, имея в виду, что система регуляции ветер представляет собой нервный способ регуляции, система желчи — гуморально-эндокринный и система слизи — местно-тканевой уровни регуляции состояния организма. Литература Ажипа А.П. — В кн.: Физиология эндокринной системы. Л., 1979. С. 535-537. Ашмарин И.П. — Журнал эволюционной биохимии и физиологии, 1979. Т. 15. С. 278—282. Ашмарин И.П. — Журнал эволюционной биохимии и физиологии, 1982. Т. 18. С. 3—10. Баклаваджян О.Г. — В кн.: Центральные механизмы вегетативной нервной системы. Ереван, 1969. С. 41—50. Балаболкин М.И. — Эндокринология. М.: Медицина, 1989. 410 с. Баранова Н.Ф. — В кн.: Физиология эндокринной системы. Л., 1979. С. 54—59. Богач П.Г. — В кн.: Физиология пищеварения. Л., 1977. С. 277-310. Богач П.Г. — Взаимоотношения гипоталамуса, лимбической системы и эндокринных желез в регуляции вегетативных функций. Киев, 1982. 135 с. Ивашкин В.Т. — Метаболическая организация функций желудка. Л.: Наука. 1981, 196 с. Климов П.К. Пептиды и пищеварительная система. Л., 1983. 271 с. Климов П.К. — Физиологическое значение пептидов мозга для деятельности пищеварительной системы. Л., 1986. 218 с. Константинова М.С. — В кн.: Физиология эндокринной системы. Л., 1979. С. 43—53. Мусящикова С.С., Черниговский В.Н. — Кортикальное и субкортикальное представительство висцеральных систем. Л., 1973..286 с. Нрздрачев А.Д. — Физиология вегетативной нервной системы. Л., 1983. 296 с. Соловьева ИА. — Архив анатомии, гистологии и эмбриологии. 1981. Т. 80. С. 88—101. Черкес В.О. — Физиол. журн. СССР. 1969. Т. 55. С. 556-565. Bloom S.R., Polak F.M. — Lancet. 1978. V. 1. P. 93. Burnstock G. — Pharmacol. Rev. 1972. Vol. 24. P. 509—580. Burnstock G. — In: Gut hormons /Ed. S.R.Bloom, FMJolak. Edinburg etc., 1981. P. 482-486. Dale — цитировано по книге П.К.Климова «Физиологическое значение пептидов мозга для деятельности пищеварительной системы. Л., 1986. 218 с. Fujita Т. — In: Chromaffin enterochromaffin and related cells /Eds.: R.E.Coupland, T.Fujita. Amsterdam, 1976. P. 191— 208. Fujita T. — Concept of paraneurons. — Arch.Histol.Iap., 1977. Vol. 40, suppl. 1. P. 1-12. Fujita Т., Kobayashi S. — In: Gastro-enteropancreatic endocrine system: A cell-biological approach /Ed.: T. Fujita, Tokyo, 1973. P. 1-16. Fujita Т., Kobayashi S., Fujii Sh., Ivanada Т., Serizawa I. — In: Cellular basis of chemical messengers hi the digestive system / Eds.: MJ.Grossman et al. New York etc., 1981. P. 231—242. Gamse R., Lembeck F., Guello S. — Naunyn-Schmiedebergs Arch. Pharmacol., 1979. Vol. 306. P. 37—44. Gersch M., Richter K. — Das peptiderge Neuron. lena, 1981. 356 p. Grossman M.I. — In: Gastrins and the vagus / Ed.: JJF.Renfeld. London, New York, 1979, P. 105—113. Grube U., Forssman W.G. — Horm. Metabol. Res., 1979. Vol. 11. P. 589-606. Hokfelt Т., Schultzberg M., Johansson O., Ljungdahl A., Elfvin L., Elde R., Terenius L., Nilsson G., Said S., Goldstein M. — In: Gut hormones / Ed. S.R.Bloom, J.M.Polak. Edinburg etc., 1978a. P. 423-433. Hokfelt Т., Ljungdahl A., Steinbusch H., Ulrhofstad A., Nilsson G., Brodin E., Pernow В., Goldstein M. — Neuroscience. 1978b. Vol. 3. P. 517-538. Humphrey C.S., Fischer J?. - Lancet. 1978. Vol. 1. P. 390. Jones C.G. — Lancet. 1978. Vol. 1. P. 272-273. Krisch В., Leenhardt H. - Cell Tiss. Res., 1980. Vol. 211. P. 251-263. Larsson L.I. — JJffistochem. Cytochem., 1980. Vol. 28. Lundberg JM., Hokfelt T, Kewenter J., Pettersson G., Ahlman H., Edin R., Dahlstrom A., Nilsson G., Terenius L., Uvnas-Wallenstein K., Said S.I. — Gastroenterology. 1979. Vol. 77. P. 468-471. Nicoll RA. — In: Nenrotransmitters; Hormones and receptors novel approaches / Ed. JAJFerendelli. New York, 1976. P. 99-122. Otsuka M., Konishi S. — Trends Neurosci. 1983. VoL 6. P. 317-320. Pearse A.G.E., Takor T.T. — Fed. Proc. 1979. VoL 38. P. 2288-2294. Samson WJL, Kodowski GP. — Bio Sci. 1981. Vol. 31. P. 445-448. Snyder 8Л. — The Harwey Lect. Ser. 73. New York, 1979. P. 291-314. Snyder SJL, Symantow R., Pasternak G.W. — In: Neurotransmitters, hormones, receptors; Novel approaches / Ed. AJerendelli. 1976. Vol. 1. P. 82-99. Stewart JM., Channabasavian K. — FedJProc., 1979. Vol. 38. P. 2302-2308. Дата добавления: 2015-12-16 | Просмотры: 574 | Нарушение авторских прав |