|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
ПОНЯТИЕ О ГЕНОТИПЕ И ФЕНОТИПЕ МИКРООРГАНИЗМОВГенотип — совокупность генов бактериальной клетки; фенотип — совокупность всех признаков и свойств, проявляемых данной культурой. В отличие от особей высших организмов, у которых исследуются признаки и свойства каждой особи, у микроорганизмов изучаются признаки и свойства в целом всей культуры (штамма), т. е. совокупности клеток, включающих миллионы и миллиарды особей. Культуры микробов могут отличаться морфологическими, физиологическими и биохимическими признаками. К морфологическим признакам относятся окраска, размер, форма, характер края и поверхности отдельно растущих колоний и т. д.; к физиологическим и биохимическим — способность или неспособность расти при пониженной или повышенной температуре, устойчивость к антибиотикам, различным ядам, облучению, отношение к питательным средам. П<* способу питания бактерии делятся на прототрофные и аук-сотрофные. Прототрофные могут жить на минимальной питательной среде (содержащей минеральные соли и углеводы) и необходимые им вещества способны синтезировать сами. В то же время получено много штаммов грибов и бактерий, которые в отличие от исходного штамма (так называемого дикого типа) лишены способности синтезировать одну или несколько аминокислот или другие факторы роста из более простых предшественников. Такие штаммы называют ауксотрофными. Ауксотрофный штамм можно идентифицировать по его неспособности расти и размножаться на синтетической среде, которая не содержит какого-то специфического фактора роста. Но если в синтетическую среду добавить компонент, который ауксотрофный штамм не способен самостоятельно синтезировать, то он начинает расти так, что его уже невозможно отличить от прототрофного. Таким образом, культура микроорганизма может быть изучена в отношении многих признаков. Гены, ответственные за синтез определенного соединения, обозначают тремя строчными буквами, соответствующими начальным буквам этого соединения. Гены исходного дикого типа обозначают со знаком «+», например, his"1" — гистидиновый ген, leu+ — лейциновый ген, arg+ — аргининовый ген и т. д. Гены чувствительности или резистентности к лекарственным препаратам, фагам и ядам обозначают буквами s (sensitive — чувствительный) и г (resistant — резистентный). Например, чувствительность к стрептомицину обозначают sti*, а резистентность — strr. Фенотип бактерий обозначают теми же символами, что и генотип, но с прописной буквы. Так, генотипам his+, leu"1", aig+, strr соответствуют фенотипы His+, Leu+, Arg+, Strr. Символы His"1", Leu+, Arg+ указывают на способность синтезировать гистидин, лейцин, аргинин, а символ Strr — на резистентность к стрептомицину. Альтернативные им признаки обозначают символами His~~, Leu~, Arg~, Str5. Генотип микроорганизмов представлен совокупностью генов, обусловливающих потенциальную возможность формирования любого их признака. Но формирование признака происходит в определенных условиях окружающей среды, которые не всегда способствуют проявлению генотипа. Так, два штамма Е. coli с генотипами lac* и 1ас~ на среде с лактозой будут иметь разные фенотипы: бактерии с генотипом 1ас+ образуют колонии красного цвета, а бактерии с генотипом 1ас~ образуют бесцветные колонии, так как они не ферментируют лактозу. При выращивании этих штаммов на среде без лактозы фенотип их будет одинаковым. Патогенный генотип одного штамма бактерий можно отличить от другого непатогенного штамма только при заражении восприимчивого животного. В организме невосприимчивого животного генотип патогенного штамма не появится. Исследования по генетике микроорганизмов показывают, что им присуща большая изменчивость. Изменения, возникающие под влиянием окружающей среды и не сохраняющиеся при переносе клеток в исходные условия, носят название модификацион-ных. Модификации не наследственны, они не затрагивают генотипа микроба и исчезают в первом или последующих поколениях. Микроорганизмам, как и всем прочим живым организмам, свойственно проявление комбинативной изменчивости, которая является наследственной. В силу того что прокариоты имеют гаплоидный набор хромосом и им несвойственно половое размножение, генетические рекомбинации у них имеют свои особенности. Рекомбинация у бактерий происходит путем переноса генетического материала из клетки донора в клетку реципиента при помощи конъюгации, трансдукции и трансформации. Эти процессы отличаются главным образом способом переноса генетического материала. КОНЪЮГАЦИЯ Конъюгация — перенос генетического материала от одной бактериальной клетки (донора) к другой (реципиенту) при их непосредственном контакте. Процесс конъюгации у бактерий обнаружили Дж. Ледерберг и Э. Татум в 1946 г. Они провели следующий эксперимент. Были отобраны два ауксотрофных мутантных штамма Е. coli К-12: не способный синтезировать метионин и биотин штамм Met~ Bio~ и не способный синтезировать треонин и лейцин штамм Thr~ Leu~. Оба штамма в течение ночи выращивали вместе на полноценной среде. Затем смешанную культуру центрифугировали, отмывали от полноценной среды и высевали на минимальную питательную среду. На минимальной питательной среде без метионина, биотина, треонина и лейцина появились прототрофные колонии Met+ Bio+ Thr+ Leu+ с частотой около 1 на каждые 107 клеток. Дополнительные опыты показали, что ни трансформации, ни трансдукции в данном случае не происходило. Из этого следовало, что образование рекомбинант-ных геномов происходило в результате контакта родительских клеток. Вскоре были получены микрофотографии конъюгирую-щих бактерий кишечной палочки, которые свидетельствовали о том, что между бактериями при конъюгации образуется цито-плазматический мостик. В 1952 г. Хейс установил неравноценную роль родительских штаммов при конъюгации. Выяснилось, что один штамм является донором (мужским), другой — реципиентом (женским). Клетки-доноры обладают половым фактором F. Он является конъюгатив-ной плазмидой и представляет собой циркулярно замкнутую молекулу ДНК. Половой фактор F автономно существует в цитоплазме. Бактериальные клетки с фактором F обозначают F"1", а не имеющие его — F~~. В составе генома конъюгативной плазмиды имеется tra-оперон, гены которого контролируют образование половых ворсинок (пилей) донорской клетки, необходимых для осуществления контакта с реципиентной клеткой, конъюга-тивнйй перенос собственной плазмиды или хромосомной ДНК, а также репликацию автономной плазмиды. Механизм переноса генетического материала при конъюгации из бактерии донора в бактерию реципиента показали В. Вольман и Ф. Жакоб. При конъюгации фактор F может перейти из мужской в женскую клетку и превратить ее в F4". Доноры F+ переносят довольно эффективно F-плазмиду во все клетки F~, a гены хромосомы — с низкой частотой (10~5). Половой фактор F обладает способностью включаться в геном бактерии и тогда из цитоплазматической структуры превращается в фрагмент хромосомы. Клетки, в которых возникает этот процесс, образуют Hfr-штамм. Доноры Hfr переносят бактериальную хромосому с фиксированной точки — сайта интеграции плазмиды, ориентированным образом и с высокой частотой (10—2— 10 3). Интегрированный F-фактор переносится последним. Генетическим методом идентифицировано около 25—30 сайтов интеграции фактора F в хромосому. При конъюгации клетки-доноры F* или Hfr соединяются с Клетками-реципиентами F~ при помощи конъюгационного мостика — особой протоплазматической трубки, образуемой клеткой F+. В клетке донора Hfr под влиянием фермента эндонуклеазы в точке внедрения фактора F происходит разрыв цепи ДНК. Свободный конец одной из цепей ДНК постепенно начинает передвигаться через конъюгационный мостик в клетку реципиента (F~) и сразу же достраиваться до двух-цепочной структуры. На оставшейся в клетке-доноре цепи ДНК синтезируется вторая цепь. Так как фактор F у разных штаммов Hfr включается в хромосому и разрывает ее в разных местах, переход хромосом в реци-пиентную клетку начинается с разных участков. Для переноса всей цепи ДНК в клетку реципиента требуется при 37 °С .100 мин, но конъюгационный мостик очень хрупкий, легко разрывается, и, как правило, вся цепь не успевает перейти. Поэтому # более высокой частотой передаются гены, расположенные около начальной 0-й точки хромосомы донора. Затем ДНК донора в гомологичных участках вступает в контакт с ДНК реципиента, и в результате кроссинговера некоторые участки одной цепи ДНК реципиента заменяются фрагментами ДНК донора. Искусственное прерывание конъюгации через определенное время после начала скрещивания и выявление рекомбинантов дали возможность определить последовательность перехода разных генов донора в клетку F~. На основании определения времени передвижения фрагментов разной длины из клеток Hfr в клетки F~ было установлено расстояние между генами в минутах, что позволило построить карты хромосом.
В основе построения карт хромосом лежат последовательность расположения генов в хромосоме и расстояние между ними в минутах. Вся окружность хромосомы Е. coli составляет 100 мин. К настоящему времени на карту Е. coli К-12 нанесено более 1000 генов, что составляет около 30 % ее генетической емкости (рис. 23). Иногда включенный в хромосому Hfr половой фактор освобождается и при этом (подобно профагу) может захватить с собой прилегающий к нему участок ДНК бактерии. При конъюгации половой фактор вместе с.фрагментом ДНК иногда переходит в женскую клетку, превращая ее в мужскую и передавая ей свойства, контролируемые фрагментом хромосомы донора. Процесс переноса генетической информации при помощи полового фактора называется сексдукцией. ТРАНСДУКЦИЯ Трансдущия — перенос генов из одной бактериальной клетки в другую при помощи бактериофага. Впервые это явление установили в 1952 г. Н. Зиндер и Дж. Ледерберг. Они проводили исследования на патогенных для мышей бактериях Salmonella typhimurium. Были отобраны два штамма этих бактерий: штамм 22А ауксотрофный, не способный синтезировать триптофан (Т~), и штамм 2А, способный синтезировать триптофан (Г1"). Эти штаммы засевали в U-образную трубку, разделенную внизу бактериальным фильтром (рис. 24). В одно колено трубки засевали штамм 22А (Т~), в другое — штамм 2А (Т4"). После определенного периода инкубации бактерии штамма 22А при посеве на минимальную питательную среду дали небольшое количество колоний (частота появления трансдуцированных клеток была равна 1-10—*). Это свидетельствовало о том, что некоторые клетки приобрели способность синтезировать триптофан. Каким же образом бактерии могли приобрести это свойство? Исследования
тезировать триптофан. Аналогичным образом могут быть трансдуцированы и другие признаки, в том числе способность к сбраживанию, устойчивость к антибиотикам и т. д.
Явление трансдукции установлено также у кишечной па- лочки и актиномицетов. Как правило, трансдуцируется один ген, реже два и очень редко три сцепленных гена. При переносе генетического материала заменяется участок молекулы ДНК фага. Фаг при этом теряет свой собственный фрагмент и становится дефектным. Включение генетического материала в хромосому бактерии реципиента осуществляется механизмом типа кроссинговера. Происходит обмен наследственным материалом между гомологичными участками хромосомы реципиента и материала, привнесенного фагом. Различают три вида трансдукции: общую, или неспецифическую, специфическую и абортивную. При неспецифической трансдукции в период сборки фаговых частиц в их головку вместе с фаговой ДНК может включиться любой из фрагментов ДНК пораженной бактерии. В результате в реципиентные клетки могут переноситься различные гены бактерии донора. Неспецифическую трансдукцию могут осуществлять фаги Р-1 и Р-22 у эшерихий, шигелл и сальмонелл. При специфической трансдукции профаг включается в определенное место хромосомы бактерии и трансдуцирует определенные гены, расположенные в хромосоме клетки донора рядом с профагом. Например, фаг X (лямбда) в состоянии профага всегда включается в одно и то же место в хромосоме кишечной палочки и трансдуцирует локус, обусловливающий способность к сбраживанию галактозы. При отделении профагов от ДНК хозяина прилегающие к профагу бактериальные гены вместе с ним выщепляются из состава хромосомы, а часть генов профага остается в ее составе. Частота общей трансдукции составляет от 1 на 1 млн до 1 на 100 млн. Специфическая трансдукция происходит чаще. Установлено, что фрагмент хромосомы донора, перенесенный в% клетку реципиента, не всегда включается в хромосому реципиента, а может сохраняться в цитоплазме клетки. При делении бактерий он попадает только в одну из дочерних клеток. Такое состояние получило название абортивной трансдукции. Дата добавления: 2015-12-16 | Просмотры: 1261 | Нарушение авторских прав |

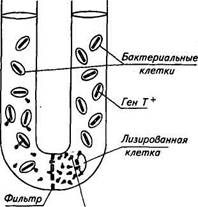 показали, что штамм 22А был лизогенен по фагу Р-22. Этот фаг освобождался из лизоген-ной культуры, проходил через фильтр и лизировал штамм 2А. Присоединив часть генетического материала штамма 2А, фаг возвращался обратно и передавал этот генетический материал штамму 22А. Штамм 22А приобретал специфические наследственные свойства штамма 2А, в данном случае свойство син-
показали, что штамм 22А был лизогенен по фагу Р-22. Этот фаг освобождался из лизоген-ной культуры, проходил через фильтр и лизировал штамм 2А. Присоединив часть генетического материала штамма 2А, фаг возвращался обратно и передавал этот генетический материал штамму 22А. Штамм 22А приобретал специфические наследственные свойства штамма 2А, в данном случае свойство син-