жду центромерой и локусом типа скрещиваемости (см. рис. 5.16). Частота возникновения таких асков составляет (9 + 5+11 + 14)/ 273 = = 0,143. В каждом аске, обнаруживающем расщепление при втором делении, имеется половина рекомбинантных и половина нерекомбинантных (родительского типа) хроматид. Таким образом, частота кроссинговера между центромерой и локусом типа скрещиваемости в расчете на хроматиду (т.е. та величина, на основании которой вычисляется расстояние по карте) составляет 1/2 · 0,143 = = 0,0715. 5.19. Наличие хроматид с незарегистрированным двойным кроссинговером увеличи-
5.17. Расщепление аллелей типов скрещиваемости при втором делении проявляют аски, в которых имел место кроссинговер ме-
вает число родительских типов и способствует занижению оценки расстояния между генами, расположенными в соответствующей области.
Глава 6
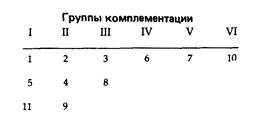
6.1. Мутанты 2 и 6 образуют одну комплементационную группу, а мутанты 1, 3 и 4 -другую. Мутант 5 не способен комплементировать мутации ни одной из этих групп и является, вероятно, делецией, инактивирующей оба гена. 6.3. Поскольку новая мутация не рекомбинирует с делецией 1241, но рекомбинирует с J3, ее можно локализовать между концами
6.9. Преобладание генотипов у/о и trp среди неселектируемых маркеров свидетельствует о том, что они фланкируют локус pan. Аллели pan можно расположить по порядку, считая, что основной генотип является результатом одиночного кроссинговера. Таким образом, порядок соответствует: уlо-рап25pan20-pan18-trp. 6.11.
этих двух делеций на участке 2 цистрона А. На участке А2 расположены концы ее шести делеций. Известно, что мутация не участвует в рекомбинации, если она находится на участке, делетированном в другой родительской хромосоме. Отсюда следует, что новая мутация расположена на участке А2с. 6.5. Показаны участки, делетированные в ДНК каждого из мутантов:
6.13. Расстояние на карте = (2-2/100000) x x 100 = 0,004 сМ. 6.15. Поскольку и рекомбинантный, и родительский генотипы соответствуют фенотипу rII, необходимо проверить генотип фагового потомства в каждой бляшке. Для этого каждый из дочерних фагов скрещивают как с r168, так и с r924. Фаги родительского типа r168 r1695 при скрещивании с r168 не будут давать рекомбинантов дикого типа, а фаги родительского типа r1695 r924 не смогут дать рекомбинантов дикого типа при скрещивании с r924. В то же время при скрещивании ре-