комбинантов r168 r1695 r924 с обоими тестерными штаммами в потомстве фаговых частиц дикого типа не будет, а при скрещивании с этими штаммами рекомбинанта r 1695
фаговые частицы дикого типа обнаружить можно. Быстрый способ скрещивания, разработанный Бензером, делает эту громоздкую процедуру достаточно простой в исполнении.
Глава 7
тифицируют классы двойных рекомбинантов и выводят взаимное расположение трех генов. На основании тех же данных рассчитывают и расстояния по карте s — co1 (2,6 ед. карты) и со1 — mi (5,4 ед. карты). В оставшихся двух скрещиваниях рассматривают mi как неселектируемый маркер, а фенотип «мутные бляшки» («+ +»), как тип селектируемых рекомбинантов. Значения частот рекомбинации в этих скрещиваниях следует умножать на 2, поскольку не учитываются реципрокные рекомбинанты (ссох).
7.15. Фаг λ имеет больше общих генов с лямбдоидным фагом 434, чем с фагами 21 и 82, при этом он более схож с фагом 21, чем с фагом 82. Ни один из трех упомянутых лямбдоидных фагов не несет гена λсI. Ген λcI обеспечивает функцию (кодирует репрессор) поддержания лизогенного состояния и обусловливает иммунность клетки к суперинфекции фагом λ, но не фагом 434. Таким образом, сайт(ы) действия репрессора cI должны располагаться между генами cII и сIII, в области, где отсутствует гомология между последовательностями ДНК фагов λ и 434. Эта область называется областью иммунитета. 7.17. Порядок маркеров: imm — 2001 — can1—42—O. 7.19. Фаг, инфицирующий Е. coli С1, но дающий потомство, неспособное реинфицировать Е. coli С1, должен содержать ДНК дикого типа, заключенную в капсид фага НаНb. Это явление носит название смешения фенотипов, поскольку в ходе инфекции фагами с двумя различными генотипами, ДНК одного из них может инкапсулироваться в белковую оболочку, синтез которой направлялся ДНК другого генотипа. Формирующиеся таким образом фаговые частицы будут обнаруживать фенотипическое проявление, нехарактерное для их истинного генотипа.

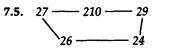
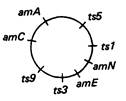

 7.15. Фаг λ имеет больше общих генов с лямбдоидным фагом 434, чем с фагами 21 и 82, при этом он более схож с фагом 21, чем с фагом 82. Ни один из трех упомянутых лямбдоидных фагов не несет гена λсI. Ген λcI обеспечивает функцию (кодирует репрессор) поддержания лизогенного состояния и обусловливает иммунность клетки к суперинфекции фагом λ, но не фагом 434. Таким образом, сайт(ы) действия репрессора cI должны располагаться между генами cII и сIII, в области, где отсутствует гомология между последовательностями ДНК фагов λ и 434. Эта область называется областью иммунитета. 7.17. Порядок маркеров: imm — 2001 — can1—42—O. 7.19. Фаг, инфицирующий Е. coli С1, но дающий потомство, неспособное реинфицировать Е. coli С1, должен содержать ДНК дикого типа, заключенную в капсид фага НаНb. Это явление носит название смешения фенотипов, поскольку в ходе инфекции фагами с двумя различными генотипами, ДНК одного из них может инкапсулироваться в белковую оболочку, синтез которой направлялся ДНК другого генотипа. Формирующиеся таким образом фаговые частицы будут обнаруживать фенотипическое проявление, нехарактерное для их истинного генотипа.
7.15. Фаг λ имеет больше общих генов с лямбдоидным фагом 434, чем с фагами 21 и 82, при этом он более схож с фагом 21, чем с фагом 82. Ни один из трех упомянутых лямбдоидных фагов не несет гена λсI. Ген λcI обеспечивает функцию (кодирует репрессор) поддержания лизогенного состояния и обусловливает иммунность клетки к суперинфекции фагом λ, но не фагом 434. Таким образом, сайт(ы) действия репрессора cI должны располагаться между генами cII и сIII, в области, где отсутствует гомология между последовательностями ДНК фагов λ и 434. Эта область называется областью иммунитета. 7.17. Порядок маркеров: imm — 2001 — can1—42—O. 7.19. Фаг, инфицирующий Е. coli С1, но дающий потомство, неспособное реинфицировать Е. coli С1, должен содержать ДНК дикого типа, заключенную в капсид фага НаНb. Это явление носит название смешения фенотипов, поскольку в ходе инфекции фагами с двумя различными генотипами, ДНК одного из них может инкапсулироваться в белковую оболочку, синтез которой направлялся ДНК другого генотипа. Формирующиеся таким образом фаговые частицы будут обнаруживать фенотипическое проявление, нехарактерное для их истинного генотипа.