Воспаление, вызываемое палочкой сапа
Воспаление, вызываемое палочкой caпa (actinobacillus mallei), отличается
разнообразием проявлений в зависимости от стадии болезни и иммунологиче-
ского состояния организма.
При о с т р о м с а п е возникают узелки, построенные из грануляционной
ткани, которая состоит из макрофагов и эпителиоидных клеток с примесью
неитрофилов. Этиузелки быстро подвергаются некрозу и гнойному распла-
влению, которое преобладает в картине болезни. Характерен распад ядер кле-
ток узелков — кариорексис: ядра превращаются в мелкие глыбки, интен-
сивно окрашивающиеся гематоксилином. В различных органах возникают
абсцессы.
Для хронического сапа характерны бугорки с преобладанием проли-
феративного воспаления, которые образуются в разных органах, в том числе
и в легких, и очень напоминают туберкулезные бугорки. В органах, в частно-
сти в легких, могут возникать склеротические изменения.
Болезни, при которых образуются гранулемы, объединены в настоящее время в большую
группу так называемых гранулематозных болезней. В их возникновении большое значение
имеет не только патогенный фактор, который может быть различным, но и иммунные нару-
шения. В последние годы отмечается учащение гранулематозных болезней в связи с увеличе-
нием в популяции лиц с повышенной реактивностью организма.
ИММУНОПАТОЛОГИЧЕСКИЕ ПРОЦЕССЫ
Иммунопатологическими называют процессы, развитие которых
связано с нарушением функции иммунокомпетентной (лимфоидной) ткани.
Иммунопатологические процессы составляют основу иммунопатоло-
гии — раздела медицины, изучающего все процессы и болезни, которые воз-
никают в результате иммунологического конфликта и наруше-
ний иммунологического гомеостаза. Помимо такого широ-
кого толкования иммунопатологии, существует другое, более узкое.
Согласно ему, под иммунопатологией понимаются аутоиммунизация,
аутоаллергия, аутоагрессия, т.е. состояния, характеризующиеся
появлением аутоантигенов и аутоантител, что ведет к повреждению клеток
и тканей организма.
Морфологическим изучением иммунопатологических процессов занимает-
ся иммуноморфологи я, которая, помимо чисто морфологических и им-
мунологических методов, широко использует иммуногистохимиче-
с к и й, или иммунолюминесцентный метод (метод люминес-
цирующих антител).
С помощью этого метода можно выявлять в тканях и клетках компоненты иммунной реак-
ции (антиген, антитело, комплемент) и устанавливать связь этой реакции с характером разви-
вающихся морфологических изменений. Нередко иммунолюминесцентный метод используется
в различных модификациях при сочетании люминесцирующих антител или антигенов с радиоак-
тивными метками и маркерами для электронной микроскопии (ферритин, ртуть, йод), а также
гистохимическими реакциями. При этом сочетание иммуногистохимического и электронно-ми-
кроскопического методов позволяет выявлять компоненты иммунного комплекса (антиген, анти-
тело, комплемент), связанные с определенными ультраструктурными элементами.
Морфология иммунопатологических процессов включает структурное вы-
ражение нарушений иммуногенеза (антигенная стимуляция или им-
мунный дефицит) и местныхиммунных реакций, совершающих-
ся в сенсибилизированном организме— реакции гиперчув-
ствительности.
МОРФОЛОГИЯ НАРУШЕНИЙ ИММУНОГЕНЕЗА
Нарушения иммуногенеза (иммунологического гомеостаза)
могут быть связаны с двумя типами иммунных реакций — гуморальной
и клеточной.
В отношении реакций гуморального или клеточного иммунитета лимфоид-
ная ткань детерминирована, т. е. разделена на лимфоциты, причастные к гу-
моральной или клеточной иммунной реакции. Реакции гуморального иммуни-
тета лимфоидная ткань осуществляет с помощью В-лимфоцитов
(бурсазависимые лимфоциты, от лат. — bursa), а клеточного имму-
нитета — с помощью Т-лимфоцитов (тимусзависимые лимфо-
циты).
В периферической лимфоидной ткани эти лимфоциты имеют свои зоны
распределения. Особенно четко эти зоны представлены в лимфатических узлах
и селезенке. В-зависимы ми зонами являются корковый слой, светлые
центры фолликулов и мозговой слой лимфатических узлов, а также перифери-
ческая зона фолликулов селезенки. К Т- зависимым зонам относятся
околокорковый слой, периферическая зона фолликулов лимфатических узлов
и околоартериальная зона фолликулов селезенки.
Имеются различия между В- и Т-лимфоцитами. В-зави-
симые лимфоциты фило- и онтогенетически более молодые, происходят
из стволовых клеток костного мозга или лимфоидных образований кишечни-
ка. Эти «оседлые» клетки имеют характерную структуру, живут сравнительно
недолго, способны при антигенной стимуляции трансформироваться в плаз-
мобласты и плазматические клетки, продуцирующие специфические иммуно-
глобулины, т. е. антитела.
Т-зависимыелимфоциты имеют тимусное происхождение, поэтому
фило- и онтогенетически они более древние. Эти долгоживущие клетки несут
на своей поверхности специфический антиген (Q-антиген), способны выпол-
нять функцию «клеток иммунологической памяти», т. е. распознавать «свое»
и «чужое». Имея свои зоны распределения в периферической лимфоидной тка-
ни, они тем не менее являются рециркулирующими лимфоцитами,
т. е. мигрируют из лимфоидных органов в кровь и обратно и «информируют»
лимфоидную ткань о состоянии иммунологического гомеостаза. Однако они
не только передают информацию об антигене, но и специфи-
чески реагируют с антигеном и способны разрушать его.
Популяция Т-лимфоцитов не является однородной. Доказано наличие Т-клеток-хелпе-
р о в (от help — помощь), Т-клеток-к и л л е р о в (от killer — убийца), Т-к лето к-с у п р е с с о-
р о в (от suppression — подавление) и Т-клеток гиперчувствительности замедлен-
ного типа (ГЗТ). Т-клетки-хелперы («помощники») кооперируют с В-лимфоцитами, что
определяет возможность их трансформации в плазматические клетки и образование антител. Т-
клетки-киллеры («убийцы») являются основными клетками-эффекторами в реакциях клеточного
иммунитета; они разрушают клетки-мишени и обеспечивают генетическое постоянство внутрен-
ней среды организма. С Т-клетками-супрессорами (блокаторами) связано развитие толерантно-
сти, так как они подавляют образование антител В-лимфоцитами. С Т-клетками связана ГЗТ.
Иммуногенез, т. е. механизм развития иммунных реакций,
определяющих иммунологический гомеостаз, имеет характерное морфологи-
ческое выражение.
В гуморальной иммунной реакции различают афферентное, цен-
тральное и эфферентное звенья. Афферентноезвено включает все этапы
от попадания в организм антигена до передачи информации о нем лимфоид-
ной ткани. Попавший в организм антиген подвергается фагоцитозу клетками
крови и ретикулоэндотелиальной системы. Однако основная роль в фагоцито-
зе антигена принадлежит макрофагам. Лишь после пассажа антигена через
макрофаг высвобождаются его детерминанты, которые опознаются Т-лимфо-
цитами-хелперами, и информация об антигене передается лимфоидной тка-
ни. Центральное звено гуморальной иммунной реакции состоит в пере-
даче рециркулирующим лимфоцитом информации об антигене тимусу,
«запоминании» тимусом этой информации и сообщении о ней В-лимфоцитам.
Посредником между антигеном и В-лимфоцитом являются те же Т-лимфо-
циты. Эфферентноезвено — пролиферация иммунокомпетентных клеток
В-зависимой лимфоидной ткани, в которых происходит гиперплазия грануляр-

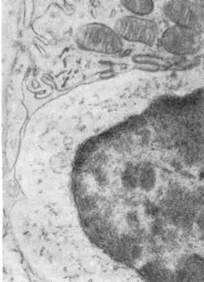
| Рис. 88. Плазмобласт селезенки,
продуцирующий антитела.
Цистерны гранулярной эндоплазма-
тической сети (ЭС) расширены, в от-
дельных участках заполнены белко-
вым содержимым (антитела); хро-
матин (Хр) в ядре расположен не-
равномерно. х 7300 (по Кингу).
ной эндоплазматической
сети, в связи с чем они
становятся более пирони-
нофильными (пирони-
нофильные клетки).
Эти клетки превращают-
ся в п л а з м о б л а с т ы
и плазма т и ч е с к и е
клетки (рис. 88), про-
исходит п л а з м а г и з а ц и я лимфоидной ткани. На высоте плазмати-
зации лимфоидной ткани отмечается наиболее высокое содержание антител
в сыворотке крови. Таким образом, гуморальная иммунная реакция направле-
на на образование специфических антител. Они способны связы-
вать антиген (реакция антиген — антитело) и образовывать иммунные ком-
плексы, которые подвергаются фагоцитозу макрофагами (иммунный фагоци-
тоз) и элиминируются различными путями.
При антигенной стимуляции антитела и иммунные комплексы циркули-
руют в избытке антигена и связывают комплемент. Такие антитела и им-
мунные комплексы повреждают клетки и ткани, в результате чего развивается
иммунное воспаление (см. с. 123).
Клеточная иммунная реакция, как и гуморальная, состоит из
трех фаз. В первую фазу (к концу суток) контакт лимфоцитов с антиге-
ном, который наблюдается в коже, слизистых оболочках, внутренних органах,
ведет к сенсибилизации Т-лимфоцитов — появляются так называемые сен-
сибилизированные (иммунные) лимфоциты (рис. 89), которые
передают информацию об антигене лимфоидной ткани. Во вторую фазу
происходит пролиферация и бластная трансформация Т-лимфо-
цитов. Образующиеся при этом бласты отличаются от плазмобластов своей
ультраструктурой и гистоферментативными свойствами (высокая активность
лизосомных фосфатаз), они не способны вырабатывать антитела. Третья
фаза состоит в реакции сенсибилизированного лимфоцита (Т-лимфоцита-
киллера) с антигеном, который чаще имеет клеточное (тканевое) происхожде-
ние (клетка-мишень). В этой реакции лимфоцит осуществляет свое цитопа-
тическое действие на клетку-мишень (рис. 90). Взаимодействие
с антигеном сенсибилизированные лимфоциты осуществляют с помощью
фиксированных на их поверхности антител (цитофильные антитела,
или рецепторы лимфоцита). Цитофильные антитела способны специ-
фически взаимодействовать с антигеном (клеткой-мишенью), при этом сенси-
билизированные лимфоциты высвобождают лизосомные ферменты, разру-
шающие антиген. При взаимодействии сенсибилизированных лимфоцитов
с антигеном выделяются также медиаторы клеточного иммуните-
та (лимфокины), с помощью которых в процесс разрушения антигена вовле-
каются макрофаги, осуществляющие иммунный фагоцитоз. Поэтому
в клеточной реакции с антигеном, помимо лимфоцита, участвуют и макрофа-
ги, причем между лимфоцитами и макрофагами появляются цитоплазма-
тические мостики (рис. 91), возникают тесные контакты.

|
Рис. 89. Имунный лимфоцит.
В цитоплазме большое число лизосом (Лз), крупные митохондрии (М), канальцы гранулярной
эндоплазматической сети (ЭС); в ядре (Я) — краевое расположение хроматина, х 23 000 (по Вильнеру).
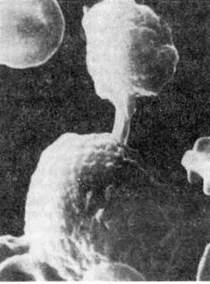
Рис. 90. Цитопатическое действие Т-лимфоцита (киллера) на клетку-мишень, х 10000.
Рис. 91. Цитоплазматический мостик между активированным иммунным лимфоцитом
(вверху) и макрофагом (внизу). Растровый электронный микроскоп, х 5750 (по Кларку и др.).
Таким образом, клеточная иммунная реакция направлена на разрушение
антигена сенсибилизированным лимфоцитом с помощью
макрофага. При антигенной стимуляции эта реакция завершается повреж-
дением клеток (иммунный цитолиз) и развитием иммунного
воспаления (см. с. 123).
Гуморальная и клеточная иммунные реакции тесно связаны и дополняют друг друга. Связы-
вающим клеточным звеном этих реакций является макрофаг, участвующий в афферентном
звене гуморальной реакции, в иммунном фагоцитозе при клеточной реакции, а также Т-лимфо-
циты-хелперы, кооперирующие с В-лимфоцитами. Связывающим гуморальным звеном можно
считать цитофильные антитела. Фиксируясь на поверхности макрофагов, они определяют их
специфическое взаимодействие с антигеном в последнюю фазу клеточной иммунной реакции.
Филогенетически реакции клеточного иммунитета являются более древни-
ми. Поэтому при внутриутробном развитии они появляются первыми. Снача-
ла возникает способность к фагоцитозу, осуществляемому стволовыми мезен-
химальными клетками. Она сменяется миелоэритроцитарным ответом,
и только к I — II месяцу внутриутробной жизни возникает лимфоидно-плазмо-
цитарный ответ.
При нарушениях иммуногенеза в связи с длительной антигенной стимуля-
цией или наследственной недостаточностью (иммунодефицитные синдромы)
возникают характерные изменения как в центральной, так и в периферической
лимфоидной ткани.
Изменения тимуса, возникающие при нарушениях иммуногенеза
Регулирующая и «цензорная» роль тимуса в иммуногенезе связана с со-
стоянием его ретикулоэпителия и лимфоцитов. Клетки тимичес-
ких, или гассалевых, телец вырабатывают гуморальный фактор тимуса, обус-
ловливающий иммунную компетенцию лимфоидной ткани.
Известно, что тимус подвергается физиологической инволюции с замеще-
нием железистой ткани жировой клетчаткой. Однако это замещение никогда
не бывает полным. Даже в старческом возрасте среди жировой ткани обнару-
живаются островки ретикулоэпителия и лимфоцитов.
Изменения тимуса, возникающие при нарушениях иммуногенеза, бывают
следующими: 1) акцидентальная инволюция (трансформация); 2) гиперпла-
зия; 3) гипоплазия. В одних случаях эти изменения являются следствием,
в других — причиной нарушений иммунологического гомеостаза.
1. При акцидентальной (от лат. accidentis — случайность) инволю-
ции (трансформации) тимуса происходит быстрое уменьшение его
массы и объема в связи с прогрессирующим распадом лимфоцитов и фагоци-
тозом их макрофагами. Гибель лимфоцитов ведет к коллапсу ретикулоэпите-
лия железы. В далеко зашедших случаях развивается атрофия тимуса.
Акцидентальная инволюция встречается у детей при многих заболеваниях,
причем степень ее тем больше, чем длительнее и тяжелее течет заболевание.
О значении акцидентальной инволюции в нарушениях иммунологического го-
меостаза нет единого мнения. Вероятно, повреждение тимуса наступает вто-
рично в связи с массивной и длительной антигенной стимуляцией. Но воз-
можно и участие нуклеопротеидов распадающихся тимоцитов в информации
лимфоидной ткани о состоянии иммуногенеза.
2. Гиперплазия тимуса нередко сочетается с гиперплазией лим-
фоидной ткани, гипоплазией надпочечников и половых желез, ожирением, уз-
кими аортой и артериями. Комплекс этих изменений, характеризующих опре-
деленный тип конституции, получил название тимик о-лимфатическо-
го состояния (status thymico-lymphaticus).
При тимико-лимфатическом состоянии в связи с дисфункцией тимуса
процессы иммуногенеза нарушены. Этими нарушениями прежде
всего объясняется чрезвычайная чувствительность людей с тимико-лимфати-
ческим состоянием к антигенной стимуляции (иммунизация, инфекционное за-
болевание), хирургическим вмешательствам, наркозу, медикаментозным пре-
паратам. От незначительных воздействий различных факторов у таких людей
может наступить внезапная смерть. Значение нарушений иммунологиче-
ского гомеостаза в развитии внезапной смерти при тимико-лимфатическом
состоянии можно объяснить тем, что тимус и лимфоидная ткань являются эф-
фекторами коры надпочечников, которая в таких случаях оказывается функ-
ционально недостаточной.
3. Гипоплазия тимуса характеризуется отсутствием деления на кор-
ковое и мозговое вещество. Ретикулоэпителий отсутствует или образует не-
многочисленные тимические тельца; лимфоцитов крайне мало. Гипоплазия
тимуса, нередко сочетающаяся с недоразвитием периферической лимфоидной
ткани, типична для комбинированного иммунного дефицита и дефицита кле-
точного иммунитета (см. с. 156).
Изменения периферической лимфоидной ткани,
возникающие при нарушениях иммуногенеза
Наиболее характерны изменения периферической лимфоидной ткани при
антигенной стимуляции и наследственной ее недостаточности.
1. При антигенной стимуляции (сенсибилизации) организма изме-
нения периферической лимфоидной ткани однозначны и выражаются м а -
крофагальной реакцией, гиперплазией ретикулярных кле-
ток и лимфоцитов с последующей плазмоцитарной их
трансформацией. Эти изменения дополняются повышением проницае-
мости микрососудов, отеком интерстиция и накоплением в нем белково-поли-
сахаридных (ШИК-положительных) веществ (тканевой диспротеиноз). Степень
макрофагально-плазмоцитарной трансформации лимфоидной ткани отражает
напряженность иммуногенеза и прежде всего уровень выработки ан-
тител (иммуноглобулинов) клетками плазмоцитарного ряда.
Особенно ярко изменения при антигенной стимуляции проявляются
в лимфатических узлах (прежде всего регионарных к месту поступле-
ния антигена) и селезенке.
В лимфатических узлах (рис. 92), которые увеличиваются, стано-
вятся полнокровными и отечными, в корковом их слое, в светлых центрах
фолликулов и мозговом слое появляет-
ся большое число плазмобластов и
плазматических клеток. Они вытесняют
лимфоциты. Отмечаются пролифера-
ция и десквамация клеток синусов, об-
разование большого количества макро-
фагов и белково-полисахаридных ве-
ществ в строме. Селезенка (рис. 93)
увеличивается, выглядит полнокровной
и сочной, на ее разрезе хорошо видны
большие фолликулы. Отмечаются рети-
кулярная гиперплазия и плазматизация
как красной пульпы, так и особенно
фолликулов селезенки, периферическая
зона которых сплошь состоит из плаз-
мобластов и плазматических клеток.
В красной пульпе наряду с плазмо-
бластами много макрофагов.
| Рис.92. Ретикулоплазмоцитарная трансфор- мация ткани лимфатического узла при анти- генной стимуляции.
| Если в ответ на антигенную стимуляцию
развиваются преимущественно клеточные им-
мунные реакции, то в лимфатических узлах и
селезенке пролиферируют в основном сенсибили-
Рис. 93. Ретикулоплазмоцитарная трансфер- Рис. 94. Макрофагально-плазмоцитарная ин-
мация фолликулов селезенки при антигенной фильтрация легких при антигенной стиму-
стимуляции. ляции.
зированные лимфоциты, а не плазмобласты и плазматические клетки. При этом происходит рас-
ширение Т-зависимых зон.
Те же изменения в виде ретикулярной гиперплазии и макрофагально-плаз-
моцитарной трансформации, а в ряде случаев и миелоидной метаплазии обна-
руживаются в костном мозге, портальных трактах и синусоидах пе-
чени, в альвеолярных перегородках, периваскулярной и перибронхиальной
ткани легких (рис. 94), в интерстиции почек поджелудочной же-
лезы, кишечника, в межмышечных прослойках, жировой
ткани и т. д.
2. Наследственная недостаточность периферической лимфоид-
ной ткани характеризуется изменениями как селезенки, так и особенно лимфа-
тических узлов. Вселезенке размеры фолликулов значительно уменьшены,
светлые центры и плазматические клетке отсутствуют. Влимфатических
узлах отсутствуют фолликулы и коркввыи слой (В-зависимые зоны), сохра-
нен лишь околокорковый слой (Т-зависимая зона). Эти изменения характерны
для наследственных иммунодефицитных синдромов, связанных с дефектом гу-
морального иммунитета (см. с. 156).
Дата добавления: 2015-11-26 | Просмотры: 607 | Нарушение авторских прав
|