|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
КЛЕТКИ ИММУННОЙ СИСТЕМЫТ-лимфоциты (рис. 4.1). Происходят Т-лимфоциты из поли-потентных костномозговых клеток. Дифференциация стволовых клеток в Т-лимфоциты осуществляется под влиянием тимозина, тимостимулина, тимопоэтинов и других гормонов, продуцируемых эпителиальными клетками или тельцами Гассаля. В процессе созревания у пре-Т-лимфоцитов (претимических лимфоцитов) постадийно реорганизуется антигенная структура цитомембран. Заканчивается дифференциация появлением у зрелых Т-лимфоцитов специфического рецепторного аппарата распознавания антигенов. Образовавшиеся Т-лимфоциты через лимфу и кровь колонизируют тимусзависимые паракортикальные зоны лимфатических узлов или соответствующие зоны лимфоидных фолликулов селезенки. Популяция Т-лимфоцитов весьма многочисленна. Их количество в периферической крови колеблется от 60 до 80 % общей численности лимфоидных клеток. Им принадлежит ведущая роль в трансплантационном, противоопухолевом и противовирусном иммунитете, гиперчувствительности замедленного типа и в аутоиммунных процессах. Зрелые Т-лимфоциты по форме напоминают малые лимфоциты крови, их ядро подковообразное, плотное и интенсивно окрашенное, цитоплазма в виде узкого ободка, диаметр 6,0...6,5 мкм. Т-клетки подразделяют на ряд следующих субпопуляций. Т-хелперы. Эти клетки генетически запрограммированы «помогать», т.е. индуцировать размножение и (или) дифференци-ровку клеток других типов. Их называют Т-хелперами и Т-индук-
ОРГАНЫ ИММУНИТЕТА И ИММУНОКОМПЕТЕНТНЫЕ КЛЕТКИ
торами. Например, они индуцируют секрецию антител В-клет-ками и стимулируют моноциты, тучные клетки и предшественников Т-киллеров к участию в клеточных иммунных реакциях. Т-хелперы активируют эффекторные, а Т-индукторы — регуля-торные клетки. Существует как минимум три типа Т-хелперов: узнающие антиген главного комплекса гистосовместимости; узнающие иммуноглобулины; секретирующие лимфокины. Часть Т-клеток осуществляет свою «хелперную» функцию через освобождение «дальнодействующих» факторов — интерлей-кина-2 и фактора, замещающего Т-клетки. Эти факторы называются лимфокинами. Они активируют или способствуют активации других Т- или В-клеток. Эти же клетки могут индуцировать реакции гиперчувствительности замедленного типа. Т-и н д у к т о р ы супрессии — это Т-клетки, индуцирующие превращение других Т-клеток в Т-супрессоры. Супрессорные Т-клетки — клетки, генетически запрограммированные для супрессорной активности. Утрата супрессор-ной активности наблюдается при ревматоидном артрите, системной красной волчанке и других аутоиммунных процессах. При повышении супрессорной активности отмечено угнетение процесса созревания иммунокомпетентных клеток, что приводит к развитию иммунодефицита. Временное повышение активности Т-супрессоров наблюдается при некоторых вирусных заболеваниях, а также хронических бактериальных инфекциях. Цитотоксические Т-клетки, или Т-киллеры. Киллер-ные клетки формируются после экспозиции с аллогенными, опухолевыми клетками или вирусами. Активность Т-киллеров зависит от функции Т-индукторов. Эта популяция Т-клеток продуцирует растворимый фактор, необходимый для дифференцировки Т-киллеров. При повторном контакте с аллогенными или опухолевыми клетками, теперь уже клетками-мишенями, киллеры при близком контакте с последними освобождают цитотоксические лимфокины, называемые лимфотоксинами, которые нарушают мембранную проницаемость клеток-мишеней и вызывают их гибель. В-клетки. Происходят В-клетки от стволовых клеток. Созревают поэтапно — первоначально в костном мозге, затем в селезенке. Предшественники В-лимфоцитов (пре-В-лимфоциты появляются на 16-е сутки внутриутробного развития плода). На самой ранней стадии созревания на цитоплазматической мембране В-клеток экс-прессируются иммуноглобулины М, несколько позже в комплексе с ними появляются иммуноглобулины G или А, а к моменту рождения, когда происходит полное созревание В-лимфоцитов, — иммуноглобулины D. В-лимфоциты являются предшественниками антителообразующих клеток. Так как большинство антигенов тимусзависимые, то для трансформации В-лимфоцитов в антитело-
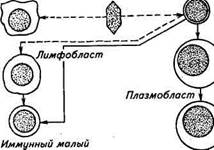
продуцирующие обычно недостаточно одного антигенного стимула. При попадании таких антигенов В-лимфоциты дифференцируются в плазмоциты с помощью Т-хелперов при участии макрофагов и стромальных ретикулярных отросчатых клеток. При этом хелперы выделяют цитокины — гуморальные эффекторы, которые и активируют пролиферацию В-лимфоцитов. Встречаются, однако, и Т-независимые антигены, такие, как, например, бактериальные липополисахариды, полисахарид стрептококка. Полимеризо-ванные белки жгутиков могут стимулировать антителообразова-ние без Т-хелперов. Правда, иммунный ответ на Т-независимые антигены ограничивается продукцией иммуноглобулинов М, и повторное их воздействие на организм не сопровождается, как обычно, усиленной выработкой антител. Пул В-лимфоцитов в организме значительный. Больше всего их в групповых лимфатических фолликулах, костном мозге, крови и селезенке (40...60 %), в лимфатических узлах и грудном лимфатическом протоке (25 %). Практически нет В-лимфоцитов только в тимусе. Диаметр зрелых В-лимфоцитов 8,5 мкм. Ободок цитоплазмы шире, а ядро светлее, чем у Т-лимфоцитов. Поверхность В-лимфоцитов покрыта густым слоем отростков — антигенрас-познающих рецепторов, которые по структуре представляют собой мономеры иммуноглобулинов М. Независимо от природы и силы антигена, который вызвал трансформацию В-лимфоцитов, образующиеся плазмоциты продуцируют антитела. Таким образом, антигенный стимул надо рассматривать как пусковой сигнал для выработки генетически запрограммированного синтеза антител. В-клетки периферической крови включают субпопуляции, по-разному реагирующие на сигналы Т-лимфоцитов. В-клетки I типа (менее зрелые лимфоциты) реагируют на хелперные действия Т-лимфоцитов только в процессе прямого межклеточного контакта. В-клетки II типа (более зрелые лимфоциты) чувствительны к растворимым факторам Т-клеток. Итоговой фазой В-клеточного иммунитета является постоянная продукция иммуноглобулинов-антител. Синтез иммуноглобулинов происходит в пузырьках пластинчатого аппарата Гольджи. Одновременно в организме синтезируется до 105...107 антител различной специфичности. Интактные В-лимфоциты синтезируют за 1 ч примерно 250...500 молекул IgM и большую часть их отдают в экстрацеллюлярную жидкость. После специфической антигенной стимуляции и трансформации лимфоцитов в плазматические клетки этот синтез возрастает в тысячи раз (рис. 4.2) В 70-х годах внимание исследователей привлекли лимфоциты периферической крови, которые не имеют мембранных структур, типичных для Т- и В- лимфоцитов, но обладают комплементзави-симой цитотоксической активностью по отношению к клеткам-мишеням, покрытым антителами. Лимфоциты с подобной характеристикой получили название ноль(0)-лимфоцитов. В пределах лимфоцит Плазмоцит Рис. 4.2. Трансформация В-лимфоцитов в ан-тителопродуцирующие клетки фракции 0-лимфоцитов выделяют L-клетки и К-клетки. L-клетки выделены только у человека. К-л имфоциты — интактные лимфоциты, разрушающие в отсутствие комплемента клетки-мишени, покрытые антителами. Антителозависимая цитотоксичность К-клетки проявляется против некоторых опухолевых клеток, аллогенных и аутогенных лимфоцитов. Специфичность цитотоксического эффекта К-клеток в отличие от специфичности Т-киллеров определяется не их собственными функциональными свойствами, а специфичностью антител, покрывающих клетки-мишени. Антитела, обеспечивающие цито-токсическое действие К-клеток, относятся ко всем четырем субклассам IgG; из них наиболее активные IgGl, 3 и 2. Антитела класса М, А и Е с К-клетками не реагируют. К-клетки продуциру: ют в присутствии иммунных комплексов фактор, подавляющий миграцию лимфоцитов. Цитотоксический потенциал К-клеток очень высок: одна клетка может лизировать не менее 10 клеток-мишеней. После контакта с К-лимфоцитами гибель клетки-мишени происходит в течение 15 мин. Дата добавления: 2015-12-15 | Просмотры: 897 | Нарушение авторских прав |